ERİTROPOEZ
Erythropoiesis (Yun. erythros, “kırmızı” + poiesis, “yaratılış, yapım”)
Eritrosit üretiminin kemik iliğinde gerçekleştiği 19. yüzyılın sonlarında kabul görmüş, ancak bu sürecin moleküler düzenleyicileri 20. yüzyılın ortalarına dek gizemini korumuştur. 1906’da Carnot’un “hemopoetin” kavramını önermesi, 1950’lerde Reissmann ve arkadaşlarının parabiyoz deneyleriyle eritropoietinin (EPO) böbrek kaynaklı olduğunu göstermesi, ve 1985’te recombinant human EPO (rhEPO) üretimi, eritropoez araştırmalarında dönüm noktalarıdır. Günümüzde, tek hücreli RNA dizileme (scRNA-seq), kromatin immunoprecipitasyonu (ChIP-seq) ve yüksek çözünürlüklü canlı hücre görüntüleme teknikleri, terminal eritropoezin son evrelerindeki enükleasyon ve retikülosit olgunlaşması mekanizmalarını atomik düzeyde aydınlatmaktadır.
İnsan embriyogenezisinde eritropoez üç fazda gerçekleşir:
- Primer (Mezodermal/Yolk Sak) Eritropoezi: Embriyonik yaşamın 3. haftasında, yolk sak mezoderminden köken alan “primitif” eritroid hücreler üretilir. Bu hücreler nükleuslarını korur, büyüktürler ve embriyonik hemoglobin (HbE, Hb Portland, Hb Gower I/II) sentezlerler. Eritroid hücrelerin dolaşımda nükleuslu olarak dolaşması, memelilerde geçici bir embriyonik özelliktir; bu hücreler muhtemelen fetal karaciğerde enükleasyonlarını tamamlarlar.
- Definitif (Karaciğer/Splenik) Eritropoezi: Embriyonik 2.-3. aylarda, aort-gonad-mezonefros (AGM) bölgesinden göç eden definitif hematopoetik kök hücreler fetal karaciğerde kolonize olur. Burada enükleasyon yeteneğine sahip, daha küçük ve HbF (fetal hemoglobin) sentezleyen eritrositler üretilir.
- Kemik İliği Devralması: Doğum öncesi dönemde (insanda yaklaşık 7. ay), hematopoez kalıcı olarak kemik iliğine kayar. Doğumdan sonra eritropoez esas olarak kemik iliğinde gerçekleşir.
Yetişkinde, eritropoez kırmızı kemik iliğinde (medulla ossium rubra) gerçekleşir. Yaşamın ilk 5 yılında tüm kemiklerin iliği aktifken, 20 yaşından sonra eritropoez öncelikle vertebralar, sternum, kaburgalar, pelvis, skapula ve kranial kemikler gibi membranöz kemiklerle sınırlıdır. Uzun kemiklerin diafizleri yağ dokusuyla (sarı kemik iliği) değiştirilir ve eritropoetik fonksiyonunu kaybeder.
Eritropoez, hematopoetik kök hücreden (HSC) olgun eritrosite uzanan, proliferasyon, diferansiyasyon ve matürasyon aşamalarını içeren kademeli bir süreçtir.
1. Megakaryocyte-Erythroid Progenitor (MEP):
HSC’den myeloid progenitörlere ayrıldıktan sonra, megakaryosit ve eritroid hücrelere ortak kılanmış progenitördür. MEP’nin eritroid hücreye mi yoksa megakaryosite mi diferansiye olacağına, GATA-1, GATA-2, PU.1, SCL/TAL1, KLF1 ve FLI-1 gibi transkripsiyon faktörlerinin dengelesi karar verir.
2. Burst-Forming Unit-Erythroid (BFU-E):
Eritroid seriye en erken bağlanmış progenitördür. Yüksek proliferatif kapasiteye sahiptir; tek bir BFU-E, çok sayıda koloni oluşturabilir. EPO reseptörleri (EPOR) bu evrenin orta-ileri aşamalarında yüzeye çıkar. İki alt tipi vardır:
- Steady-state BFU-E: Kemik iliğinde, homeostatik üretimden sorumlu.
- Stress BFU-E: Splenik veya hepatik mikroçevrede, BMP4, SCF ve hipoksi sinyallerine yanıt veren, kendini-yenileyen (self-renewing) popülasyon.
3. Colony-Forming Unit-Erythroid (CFU-E):
BFU-E’den türeyen, daha az proliferatif ancak EPO’ya tam bağımlı progenitördür. Yüzeyde maksimum EPOR sayısına ulaşır ve EPO yokluğunda apoptozise (programlı hücre ölümüne) mahkûmdur. CFU-E evresi, terminal eritropoezin başlangıcı olarak kabul edilir.
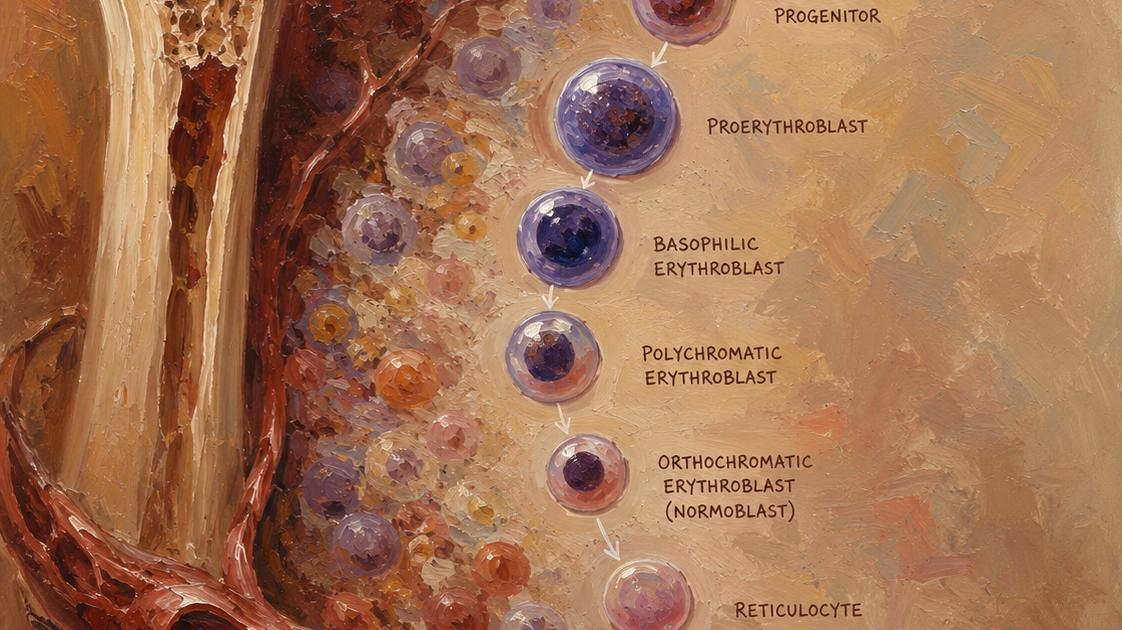
CFU-E’den sonra başlayan terminal diferansiyasyon, morfolojik olarak tanımlanabilir evreleri içerir. Bu evrelerde hücre boyutu küçülür, sitoplazma/nükleus oranı artar, kromatin giderek yoğunlaşır ve hemoglobin birikimi hızlanır.
1. Proeritroblast (Pronormoblast/Rubriblast):
- Büyük hücre (15-20 μm), yuvarlak merkezi nükleus, ince kromatin, belirgin nükleolus.
- Yüksek RNA sentezi; globin mRNA transkripsiyonu başlar.
- EPO’ya bağımlıdır.
2. Bazofilik Eritroblast (Erken Normoblast):
- Sitoplazma derin mavi-bazofilik (ribozomların yoğunluğu nedeniyle).
- Nükleus kondenzasyonu başlar; nükleolus kaybolur.
- Globin zinciri sentezi hızlanır; heme sentezi mitokondride devam eder.
- Hücre boyutu küçülmeye başlar.
3. Polikromatofilik Eritroblast (Orta Normoblast):
- Sitoplazma “polikromatofilik” görünüm kazanır: hemoglobinin pembe tonu ile ribozomların maviliği karışır.
- Nükleus belirgin şekilde kondense olur; nükleer hacim 10 kat küçülebilir.
- Globin sentezi doruk noktasına ulaşır.
- EPO bağımsızlığı kazanır; son evrelerdeki farklılaşma için diğer sitokinlerin rolü tam olarak aydınlatılmamıştır.
4. Ortokromatik Eritroblast (Geç Normoblast):
- Sitoplazma tamamen pembe-kırmızı (hemoglobin doygunluğu).
- Nükleus son derece kondense, piknotik (yoğunlaşmış) hale gelir.
- Hücre döngüsünden çıkar (G0/G1 arrest).
- Genomun büyük bölümü transkripsiyonel olarak susturulur; yalnızca globin ve eritroid membran proteinleri için transkripsiyon devam eder.
5. Enükleasyon ve Retikülosit Oluşumu:
Ortokromatik eritroblastın piknotik nükleusu sitoplazmadan dışarı atılır. Bu üç basamaklı bir süreçtir:
- Polarizasyon: Nükleus hücrenin bir ucuna göç eder.
- Ekstrüzyon: Nükleus, aktin-miyozin kontraktil halkası ve mikrotübül ağı tarafından 3-10 dakika içinde hızla dışarı itilir.
- Ayırma: Yeni oluşan retikülosit hareketli özellik gösterir; atılan nükleus kemik iliği makrofajları tarafından fagosite edilir (eferez).
6. Retikülosit:
- Nükleussuz, ancak içinde ribozomal RNA (rRNA) ve organeller (mitokondri, ribozom) içeren immatüre eritrosit.
- Kemik iliğinden perifere salınır; 1-2 gün içinde olgun eritrosite dönüşür.
- Olgunlaşma sırasında: autofaji ve ubikitin-proteazom sistemi ile mitokondri ve ribozomlar elimine edilir; membran ve proteom yeniden modellenir.
7. Olgun Eritrosit:
- 7-8 μm çapında, bikonkav disk şeklinde, nükleussuz hücre.
- Sitoplazmasının %95’i hemoglobindir.
- 120 günlük yaşam süresi sonunda splenik ve hepatik fagositozla yıkılır.

Eritropoezin ana düzenleyicisi Eritropoietin (EPO), böbrek peritübüler interstisyum hücreleri (yetişkinde %90) ve karaciğer (fetal dönemde ve yetişkinde az miktarda) tarafından, hipoksiye yanıt olarak üretilen glikoprotein bir sitokindir.
EPO/EPOR/JAK2/STAT5 Sinyal Yolu:
EPO, eritroid progenitörler yüzeyindeki EPO Reseptörü (EPOR) homodimerine bağlanır. EPOR, sitokin sınıfı I reseptör ailesine aittir. EPO bağlanması, önceden bir arada bulunan EPOR monomerlerinin konformasyonel değişimine yol açar ve JAK2 (Janus kinase 2)‘nin aktivasyonunu tetikler.
Aktive olmuş JAK2, EPOR’un sitoplazmik domainindeki tirozin rezidülerini fosforile eder. Bu fosforilasyon, çoklu sinyal kaskadlarını başlatır:
- JAK2/STAT5 Yolu (Kanonik Yol):
- JAK2, STAT5A ve STAT5B’yi (yapısal olarak %94 homoloji) fosforile eder.
- Fosforile STAT5 (pSTAT5), SH2 domainleri aracılığıyla paralel dimerler oluşturur.
- Importin α3/β1 aracılığıyla nükleusa taşınır.
- GAS motiflerine (TTCYXRGAA) bağlanarak hedef genlerin transkripsiyonunu aktive eder.
- Hedef genler arasında: Bcl-xL (anti-apoptozik), Pim1, Cish (negatif geribildirim), SOCS ailesi, Podocalyxin, Cyclinler ve eritroid transkripsiyon faktörleri (GATA1, KLF1, TAL1) bulunur.
- PI3K/AKT Yolu: Hücre sağkalımı ve metabolizma düzenlemesi.
- RAS/MAPK (ERK1/2) Yolu: Proliferasyon ve diferansiyasyon.
- PKC Yolu: Hücre farklılaşmasına katkı.
Negatif Geribildirim Mekanizmaları:
- CISH (Cytokine-inducible SH2-containing protein): STAT5 hedef genidir; EPOR’un ubikitinasyonu ve proteazomal degradasyonunu sağlar.
- SOCS1/SOCS3: JAK2 degradasyonu ve STAT5 inhibisyonu.
- SHP-1 (PTPN6): EPOR sitoplazmik domainine bağlanarak sinyali bastırır.
- LNK (SH2B3): JAK2 aktivasyonunu ve downstream sinyalleri inhibe eder.
- PIM1 Kinaz: SOCS1/SOCS3’ü fosforile-yarak stabilizasyonlarını artırır, dolaylı olarak STAT5’i inhibe eder.
Eritroid hücre kaderi, transkripsiyon faktörlerinin dinamik ağı tarafından belirlenir. ChIP-seq çalışmaları, bu faktörlerin eritroid genlerin promoter ve enhancer bölgelerinde kooperatif olarak işgal ettiğini göstermiştir.
1. GATA-1 (Eritroid Transkripsiyon Faktörü):
- Çinko parmaklı transkripsiyon faktörü; GATA motiflerine (WGATAR) bağlanır.
- Eritroid, megakaryosit ve mast hücre farklılaşmasında ana düzenleyicidir.
- GATA-1, hem aktivatör hem de represör olarak işlev görür; bu ikili rol, kofaktör bağlanması ve hücresel bağlama göre değişir.
- Hbb (β-globin), Hba (α-globin), Slc4a1 (band 3), Gypc (glikoforin C) gibi eritroid-spesifik genleri aktive eder.
- GATA-2’yi baskılayarak (GATA switch), eritroid matürasyonda kritik bir geçişi sağlar.
2. GATA-2:
- Erken hematopoetik progenitörlerde yüksek; GATA-1 arttıkça azalır.
- HSC’lerin sürdürülmesinde ve erken progenitör genişlemesinde rol alır.
- GATA-1/GATA-2 dengesi, eritroid/megakaryosit kader kararını etkiler.
3. KLF1 (Krüppel-like Factor 1 / EKLF):
- CACCC box’a bağlanan çinko parmaklı faktör.
- β-globin transkripsiyonunun ana aktive edicisidir. KLF1 yokluğunda embriyonik ölüm gerçekleşir.
- SWI/SNF kromatin remodelleme kompleksi (E-RC1) ve histon asetiltransferazlar (CBP/p300) ile etkileşerek β-globin lokusunda DNaz I hipersensitivite bölgesi oluşturur.
- LRF (Zbtb7a) genini de düzenleyerek, GATA-1 ile regülatuar döngü oluşturur.
4. FOG-1 (Friend of GATA-1 / ZFPM1):
- GATA-1’in kofaktörü; transkripsiyonel aktivasyon ve baskılanmada rol alır.
- FOG-1 olmadan GATA-1, eritroid genleri yeterince aktive edemez.
- Megakaryosit-eritroid kader kararında GATA-1/FOG-1 kompleksi kritiktir.
5. TAL1/SCL (T-cell Acute Lymphocytic Leukemia Protein 1):
- bHLH (basic helix-loop-helix) transkripsiyon faktörü.
- Hematopoetik kök hücrelerin ve eritroid progenitörlerin gelişiminde zorunludur.
- GATA-1, KLF1 ve LDB1 ile kompleks oluşturarak eritroid enhancerları işgal eder.
6. PU.1:
- Myeloid ve lenfoid farklılaşmada rol alan ETS ailesi faktör.
- Eritropoezde baskılayıcı rolü vardır; GATA-1 ile karşılıklı inhibisyon gösterir. PU.1 baskısı, eritroid kaderin kilidini açar.
7. LRF (Leukemia/Lymphoma Related Factor / Zbtb7a):
- KLF1’in hedef genlerinden biridir; aynı zamanda KLF1 promoterini GATA-1 ile birlikte işgal eder.
- Eritroid ve megakaryosit kaderi arasında dengeleyici rol oynar.
8. mikroRNA’lar:
- miR-451: Eritroid olgunlaşmada kritik; eritroid progenitörlerin proliferasyonunu kontrol eder.
- miR-150: Megakaryosit kaderini destekler; eritroid kaderi baskılar.
Terminal eritropoezde kromatin kondenzasyonu ve enükleasyon, epigenetik modülatörlerin koordineli eylemiyle gerçekleşir:
- Histon deasetilazlar (HDAC): Kromatinin yoğunlaşmasını sağlar.
- Histon metiltransferazlar (SUV420H2): Heterokromatin oluşumunu tetikler.
- Nükleer açılım ve histon salınımı: Kondenzasyon için kritik; miyelodisplastik sendromlarda (MDS) bu süreçteki defektler gözlemlenmiştir.
- DNA metilasyonu: Globin gen anahtarlamasında (hemoglobin switching) rol alır.
Hemoglobin, her biri bir globin zinciri ve bir heme grubu içeren dört polipeptid alt biriminden (tetramer) oluşur.
Globin Zincirleri:
- α-globin: Kromozom 16’da (α-globin gen kümesi: ζ, α2, α1).
- β-globin: Kromozom 11’de (β-globin gen kümesi: ε, Gγ, Aγ, ψβ, δ, β).
Gelişimsel evrelerde farklı hemoglobin tipleri üretilir:
- Hb Gower I (ζ2ε2), Hb Gower II (α2ε2), Hb Portland (ζ2γ2): Embriyonik.
- HbF (α2γ2): Fetal; oksijen afinitesi yüksek, plasental transferi optimize eder.
- HbA (α2β2): Yetişkin; toplam hemoglobinin %95-98’i.
- HbA2 (α2δ2): Yetişkin; %1-3.
Heme Sentezi:
Mitokondri ve sitozol arasında mekansal olarak bölünmüş 8 enzimatik basamak içerir. Başlangıç substratları glisin ve süksinil-KoA’dır; son ürün protoporfirin IX, Fe²⁺ ile birleşerek heme oluşturur. Heme, globin transkripsiyonunu pozitif olarak düzenler.
Normal eritropoezde α ve β globin zincirleri eşit oranda sentezlenir ve HbA tetramerlerini oluşturur. Ancak zincir dengesizliği (örneğin β-talasemide β zincir eksikliği) patolojik sonuçlar doğurur:
- α-hemoglobin stabilize proteini (AHSP): Serbest α-globin monomerlerinin katlanmasını ve agregasyonunu önler; temel ve hidrofobik (BH) domainine bağlanır.
- Ubikitin-Proteazom Sistemi (UPS): Fazla veya hatalı katlanmış globinleri elimine eder.
- UBE2O: Eritropoez sırasında indüklenen, hem E2 ubikitin konjuge enzim hem de E3 ligaz aktivitesine sahip bir faktör. Retikülosit evresinde proteomun yeniden şekillenmesinde ve fazla α-globinin proteazomal degradasyonunda hayati rol oynar.
β-talasemide, erken eritroid prekürsörlerde fazla α-globin başlangıçta proteazom tarafından elimine edilmeye çalışılır. Ancak hücre olgunlaştıkça bu kompansatuvar mekanizma yetersiz kalır; serbest α-globin agregatları toksik prensipitatlar oluşturur, protein kalite kontrol sistemlerini bozar ve eritroid prekürsörlerin erken apoptozisine (ineffective erythropoiesis) yol açar.
Enükleasyon, memelilerin evrimsel olarak kazandığı bir özelliktür ve eritrositlerin bikonkav şekil alarak gaz değişimini optimize etmelerini sağlar.
Moleküler Mekanizma:
- Kromatin kondenzasyonu: Proeritroblasttan ortokromatik eritroblasta kadar nükleer hacim 10 kat azalır. Histon modifikasyonları ve lamin A/C disasemblisi rol alır.
- Nükleer polarizasyon: Mikrotübül ağı ve kinesin/dynein motor proteinleri, piknotik nükleusu hücrenin bir ucuna çeker.
- Kontraktil halka: Aktin, miyozin II, filamin ve tropomiyozinden oluşan halka, nükleusun sitoplazmadan dışarı itilmesini sağlar.
- RhoA/ROCK sinyali: Hücre korteksindeki sinyalleşme, enükleasyonun koordinasyonunda kritiktir.
- Membran sıralanması: Lipid raftları ve adezyon molekülleri (örneğin ICAM-4), retikülosit ile atılan nükleus arasında ayrım oluşturur.
Atılan nükleus (piknotik cisim), kemik iliğindeki makrofajlar (özellikle “nurse macrophages”) tarafından eferez (fagosite edilerek yok edilme) yoluyla elimine edilir.
Enükleasyondan sonra retikülosit, dolaşıma salınmadan önce veya sonra son olgunlaşma adımlarını tamamlar:
Organell Eliminasyonu:
- Mitokondri: Mitofaji (mitokondriye özgü autofaji) yoluyla yok edilir; bu süreçte Nix (BNIP3L) reseptörü kritik rol oynar.
- Ribozomlar: Ubikitin-proteazom sistemi ve autofaji ile elimine edilir.
- Proteom Remodelleme: UBE2O aracılığıyla ~1000’den fazla proteinin seviyesi düzenlenir; retikülosit proteomu, neredeyse tamamen hemoglobin ve eritroid membran proteinlerinden oluşacak şekilde “basitleştirilir”.
Membran Remodelleme:
- Spektrin, ankyrin, aktin ve protein 4.1R’den oluşan sitoskeleton ağı yeniden organize edilir.
- Glikoforinler ve bant 3 proteinleri, membran stabilitesi ve şekil için kritik yapısal bileşenlerdir.
- Sıvı geçirgenliği ve iyon transportu (band 3, aquaporin-1) optimize edilir.
Hemoglobin Konsantrasyonu:
- Sitoplazma neredeyse tamamen hemoglobinle doldurur; bu, hücrenin oksijen taşıma kapasitesini maksimize eder.
Akut anemi (kanama, hemoliz) veya hipoksi durumlarında, steady-state eritropoezi yetersiz kalır. Bu durumda stres eritropoezi aktive olur.
Fare modellerinde stres eritropoezi dalakta (splenik kırmızı pulpa) ve karaciğerde gerçekleşir. İnsanda ekstramedüller hematopoez, ağır anemilerde karaciğer, dalak ve lenf nodlarında gözlemlenebilir.
Stres eritropoezi, steady-state progenitörlerden farklı, kendini-yenileyen stres eritroid progenitörler kullanır. Bu süreç üç ana sinyalin koordinasyonunu gerektirir:
Bone Morphogenetic Protein 4 (BMP4):
- Dalakta stres BFU-E’lerin genişlemesini tetikler.
- Hipoksi, HIF-2α aracılığıyla BMP4 gen ifadesini indükler.
- Tek başına yeterli değildir; SCF ve hipoksi ile sinerji gösterir.
Stem Cell Factor (SCF) / KIT Ligandı:
- KIT reseptörü (CD117) üzerinden etki eder.
- Stres BFU-E’lerin proliferasyonunu ve hayatta kalmasını sağlar.
- KIT reseptörünün tirozin 567 fosforilasyonu, akut anemi sonrası iyileşme için zorunludur.
Hipoksi (%1 O2):
- BMP4 ve SCF yanıtlarını potansiyelleştirir.
- Dalak ve karaciğerdeki stres progenitörlerin maksimum genişlemesi için gereklidir.
Hedgehog Sinyali:
- Dalak mikroçevresinde, kemik iliğinden göç eden progenitörlerin “stres progenitör kaderi” kazanmasını sağlar.
- Smoothened (Smo) reseptörü aracılığıyla çalışır; Hedgehog sinyali olmadan stres progenitörleri yenilenemez.
Stres eritropoezi, 45 kat artışlarla stres BFU-E popülasyonunu hızla genişletebilir ve 6-7 gün içinde anemiyi telafi edebilir.
İneffective erythropoiesis, kemik iliğinde eritroid prekürsörlerin aşırı sayıda üretilmesine rağmen, çoğunun olgun eritrosite dönüşmeden erken apoptozise uğradığı patolojik bir durumdur.
- Globin zincir dengesizliği: β-talasemide α-globin fazlalığı, eritroblastlarda toksik agregatlar oluşturur.
- Prematür PS ekspozisyonu: Fosfatidilserin (PS) hücre yüzeyine çıkar; bu, eritrositlerin makrofajlar tarafından erken fagosite edilmesine (eryptosis) yol açar.
- Demir metabolizması bozukluğu: Hepcidin baskılanması (eritroferron/ERFE aracılığıyla) ve artmış demir emilimi, sekonder demir yüklenmesine neden olur.
β-talasemide ineffective erythropoiesisi yansıtan belirteçler:
- Eritroferron (ERFE): Eritroblastlar tarafından salınan, hepcidini baskılayan hormon.
- Growth Differentiation Factor-15 (GDF-15): Eritroid stres indikatörü.
- Soluble Transferrin Receptor (sTfR): Demir alımı ve eritroid proliferasyon belirteci.
- CA15.3 ve Cell-free DNA (cfDNA): Artmış eritroid apoptozisini gösterir.
- Ferritin: Demir yüklenmesi göstergesi.
1. Demir Eksikliği Anemisi:
Heme sentezi için zorunlu demir eksikliği, hemoglobin üretimini kısıtlar. Eritroid prekürsörlerde “demir yoksunu” heme sentezi, mikrositik ve hipokromik eritrositlerle sonuçlanır.
2. B12/Folat Eksikliği (Megaloblastik Anemi):
DNA sentezi bozukluğu; eritroblastların sitoplazmik olgunlaşması nükleer olgunlaşmadan hızlıdır. Büyük, oval, nükleer dismaturite gösteren megaloblastlar oluşur; ineffective erythropoiesis eşlik eder.
3. Anemi of Kronik Hastalık (AI/ACD):
İnflamatuvar sitokinler (IL-6) hepcidini artırarak demir metabolizmasını bloke eder; eritropoez baskılanır. “Fonksiyonel demir eksikliği” tablosu oluşur.
4. Aplastik Anemi:
Kök hücre yetersizliği; tüm hematopoetik seriler etkilenir. Eritropoez ciddi şekilde baskılanır.
Ribozomopati olarak tanımlanan, konjenital eritroid hipoplazi ile karakterize nadir bir sendromdur (5-7 vaka/1 milyon canlı doğum). Patogenezi:
- Ribozomal protein (RP) gen mutasyonları: RPS19 (%25 olguda), RPS24, RPS17, RPL35a, RPL5, RPL11 ve diğerleri.
- Haploinsüfisiyens: Tek allel mutasyonu, ribozom biyogenezinde blokaja yol açar.
- Nükleoler stres ve p53 aktivasyonu: RPS19 yetersizliği, p53 stabilizasyonu ve p21 aracılığıyla G1/S faz arrestine neden olur.
- MYB ve KIT azalması: Proliferasyon sinyali zayıflar; SCF yanıtı bozulur.
- Eritroid spesifite nedeni: Eritroblastlar, terminal diferansiyasyonda transkripsiyonel susturma ve enükleasyona uğradıklarından, erken evrelerde çok yüksek ribozom sentez hızına ihtiyaç duyarlar. Bu “ribozom açlığı”, eritroid hücreleri diğer serilere göre daha hassas kılar.
Tedavide kortikosteroidler (prednizon) olguların ~2/3’ünde yanıt verir; kök hücre nakli tek kuratif tedavidir.
α-Talasemi: α-globin gen delesyonları/mutasyonları; HbH (β4) veya Hb Bart’s (γ4) oluşumu.
β-Talasemi: β-globin mutasyonları; α-globin fazlalığı, ineffective erythropoiesis, hemoliz, ekstramedüller hematopoez, splenomegali ve sekonder hemokromatozis.
JAK2 V617F mutasyonu: JAK2’nin katalitik domainindeki valin-fenilalanin değişimi, EPO bağımsız konstitutif JAK2/STAT5 aktivasyonuna yol açar. Sonuçta EPO bağımsız eritroid proliferasyon, polisitemi, tromboz ve splenomegali gelişir. Mutant JAK2’nin onkojenik aktivitesi için EPOR’un Y343, Y460 ve Y464 tirozin rezidülerinin fosforilasyonu gereklidir.
Eritroid serideki dismaturite; kromatin kondenzasyon defektleri, enükleasyon bozuklukları ve ineffective erythropoiesis eşlik eder. Ring sideroblastlar, mitokondriyal demir birikimini gösterir.
1. Recombinant Human EPO (rhEPO) ve Analogları:
- Epoetin alfa, beta, darbepoetin alfa, CERA: Kronik böbrek hastalığı anemisi, kemoterapi indükleyici anemi, prematürite anemisinde kullanılır.
- Darbepoetin alfa: Daha uzun yarı ömürlü (N-glikozilasyon artışı).
2. HIF Prolyl Hidroksilaz İnhibitörleri (HIF-PHİ):
- Roxadustat, daprodustat, vadadustat.
- HIF-1α/2α’nın proteazomal degradasyonunu engeller; endojen EPO sentezini artırır. Böbrek yetmezliğinde anemi tedavisinde oral kullanım avantajı sağlar.
3. Luspatercept:
- TGF-β süper ailesi sinyal inhibitörü; β-talasemi ve MDS’de ineffective erythropoiesisi azaltarak transfüzyon ihtiyacını düşürür.
4. İlaçlara Bağlı Eritropoez Baskılanması:
- Kloramfenikol, sülfonamidler, zidovudin (AZT): Eritroid progenitörler üzerinde toksik.
- Alkol: Folate metabolizmasını bozar; megaloblastik değişikliklere yol açar.
Eritropoez, evrimsel olarak optimize edilmiş, çok katmanlı bir biyolojik süreçtir. Hematopoetik kök hücreden olgun eritrosite uzanan bu yolculuk, transkripsiyon faktör ağlarının (GATA-1, KLF1, FOG-1), sitokin sinyalleşmesinin (EPO/EPOR/JAK2/STAT5), epigenetik modülatörlerin, protein kalite kontrol mekanizmalarının ve sitoskeleton dinamiğinin olağanüstü bir senkronizasyonunu gerektirir.
Enükleasyon, retikülosit olgunlaşması ve hemoglobinizasyon, eritrositlerin 120 günlük yaşam süreleri boyunca oksijen taşıma misyonunu eksiksiz yerine getirmelerini sağlayan moleküler ayarlamalardır. Stres eritropoezi gibi kompansatuvar mekanizmalar, vücudun oksijen homeostazını korumadaki esnekliğini gösterir.
Eritropoezin anlaşılması, anemi tedavisi (EPO replasmanı), talasemi yönetimi (transfüzyon, kelaynak, Luspatercept), miyeloproliferatif neoplazmların hedefe yönelik tedavisi (JAK2 inhibitörleri) ve gelecekteki ex vivo eritrosit üretimi (kültüre edilmiş eritrositler) gibi alanlarda kritik öneme sahiptir. Bu kapsamlı biyolojik bilgi, hem temel hematoloji bilgisini hem de klinik uygulamaların moleküler temelini oluşturur.