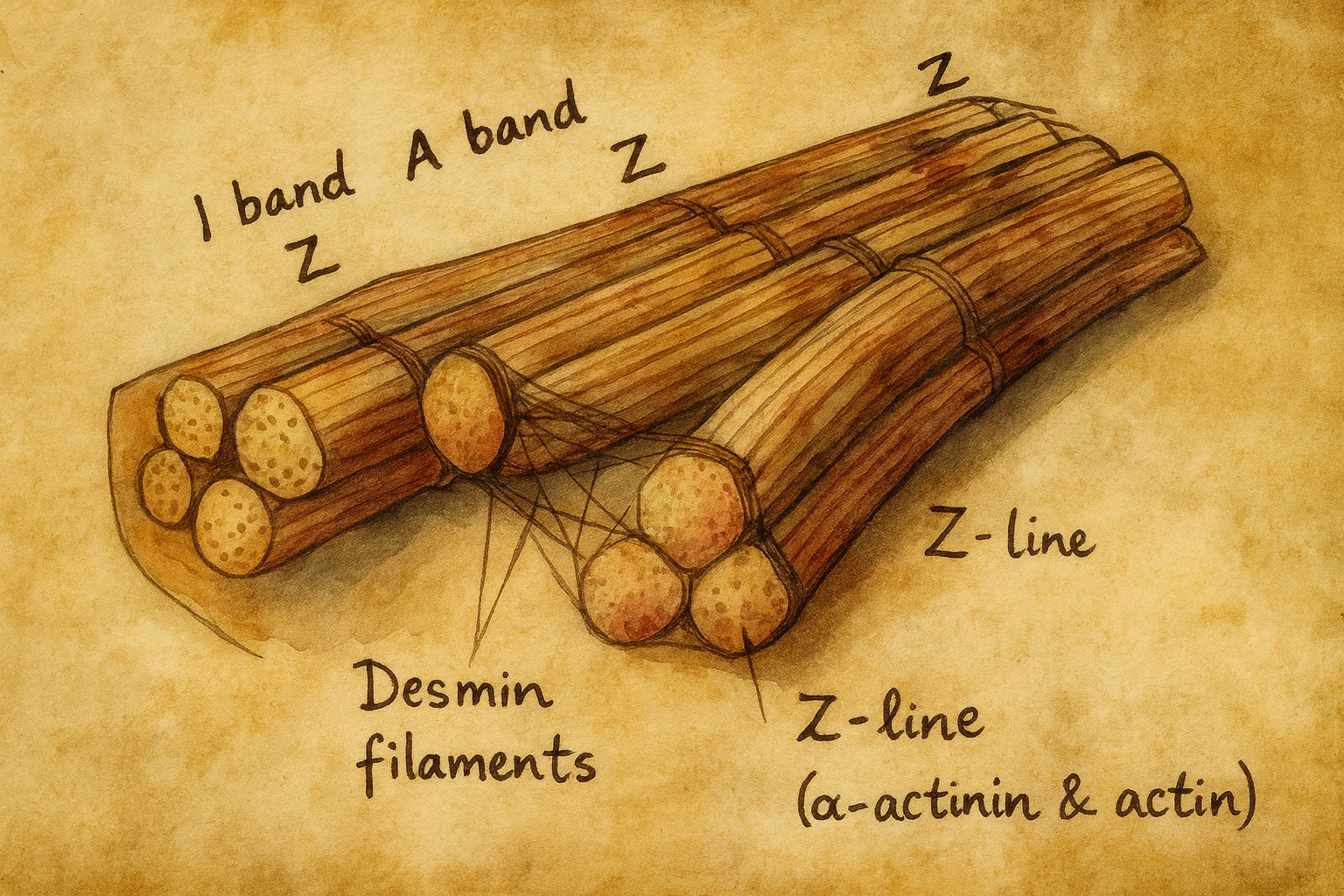
Desmin’in hikâyesi, hücre iskeletinin üçüncü kolu olan intermediyer filamentlerin kas hücresinde bıraktığı ilk izlenimle başlar. 1960’ların sonlarından itibaren elektron mikroskopisi, kas hücrelerinde 100 Å (≈10 nm) çaplı iplikçikleri yakaladığında, bu yeni fibrillerin ne olduğu tam olarak bilinmiyordu; bilinen iki “dev” vardı: mikrofilamentler (aktin) ve mikrotübüller. Kas hücresinde görülen bu üçüncü filament ailesi, bir süre “10-nm filamanlar” olarak anıldı; işlevleri belirsiz, fakat varlıkları tartışmasızdı.
1970’lerin ortasında, kas biyolojisinin deneysel dilini iyi konuşan araştırmacılar, bu iplikçiklerin kimliğini çözmek üzere kas dokusunu adeta “iskeletine” kadar soydular. 1976’da Peter Cooke, damar ve düz kas üzerinde yaptığı morfolojik çalışmalarla bu filamentöz iskeletin kas hücresindeki yaygınlığını ve mimarisini ikna edici biçimde sergiledi; kas hücresinin kontraktil aparatının yanında, ondan bağımsız ve onu tamamlayan bir “taşıyıcı ağ” vardı. Aynı yıl Elias Lazarides ve Bruce D. Hubbard, tavuk taşlığı gibi zengin düz kas kaynaklarından elde ettikleri 100 Å’lık filament altbirimini immünolojik olarak karakterize ettiler: kuvvetli üre koşullarında çözünebilen, ≈50 kDa’lık yeni bir protein, aktin ve miyozinden farklı ama kas hücresine derinden bağlı bir yapı taşı.
Bu bağ vurgusu, kısa sürede bir ada dönüştü. Yunanca δεσμός (desmos: bağ, düğüm) kökünden hareketle Lazarides ve Hubbard, proteine “desmin” adını verdiler. Aynı dönemde Joseph V. Small ve Andrzej Sobieszek, düz kastan saflaştırdıkları benzer ara filament proteinine “skeletin” adını önerdi; birkaç yıl boyunca literatürde iki isim yan yana dolaştı. Fakat “desmin”in hem biyokimyasal tutarlılığı hem de “bağ” fikrini kasın mimarisiyle özdeşleştirmesi, terimi baskın hâle getirdi. Kısacası, kavramsal yarışmayı “desmin” kazandı; çünkü isim, proteinin varlık gerekçesini anlatıyordu: sarkomerleri, z-diskleri, kostamerleri ve zarı birbirine bağlayan bir ağ kurmak.
1980’lerin başında, ara filamentlerin “üç parçalı” tasarım ilkesi —N-uç “head”, ortada uzun α-sarmal “rod”, C-uç “tail”— desmin üzerinde çözüldü. Bu yalnızca bir anatomik betimleme değildi; polimerizasyonun, yan-yana paketlenmenin ve “unit-length filament”ten olgun 10-nm filamentlere uzanan öz-montajın kuralları da belirlendi. Tripartit mimarinin keşfi, desminin neden bazen kusursuz kablolar gibi, bazen de stres altında yeniden örülen bir ağ gibi davrandığını açıklamaya başladı. Aynı yıllarda, desminin yalnızca çizgili ve kalp kasında değil, düz kasın da başat ara filament komponenti olduğu gösterildi; böylece desmin, “kas” kimliğinin moleküler amblemi hâline geldi.
1989’da insan DES geninin klonlanmasıyla öykü moleküler düzleme indi. Kod dizisi, baş-çubuk-kuyruk bölge düzenini ve koiled-koil fiziğini genetik dilden okunabilir kıldı; promotör düzenlenmesi ve miyogenez boyunca ekspresyon paternleri haritalandı. Ardından 1990’larda genetik mühendisliğin “Fonksiyon nedir?” sorusuna verdiği en net yanıtlardan biri geldi: desmin “knockout” (DES-/-) fareler. Bu hayvanlar, başlangıçta şaşırtıcı biçimde yaşayabilir ve üreyebilirdi; ancak büyüdükçe kalp ve iskelet kasında myofibrillerin yan-yana hizalanması bozuluyor, z-diskler arası eşgüdüm kayıyor, mitokondriler kümeleniyor ve kas dokusu dejenerasyona uğruyordu. “Desmin ne işe yarar?” sorusu, canlı hayvan modelinde berrak bir yanıt bulmuştu: kas dokusunda mekaniği süreklileştiren, kuvveti sarkomerden zara ve çekirdeğe köprüleyen, organellerin konumunu ve dayanıklılığını belirleyen bir “bağ örgüsü”.
İnsanda ilk desminopati olguları 1990’ların sonuna doğru tanımlandığında, temel bilim ve klinik patoloji nihayet aynı cümlede buluştu. DES genindeki mutasyonların, iskelet ve kalp kasını birlikte hedef alan myofibriller miyopatilerden ileti bozukluklu kardiyomiyopatilere uzanan geniş bir fenotip ürettiği gösterildi. Kas biyopsilerinde desminin birikim ve agregasyon paterni, ışık ve elektron mikroskopisinde z-disk düzeyindeki düzensizliklerle örtüşüyordu. Kader ortağı şaperon proteini αB-kristalinle ilişkisi giderek önem kazandı: desmin ağının sağlam kalması yalnızca polimer fizikleriyle değil, stres proteinleri ve proteostazla da ilgiliydi.
2000’lerde hikâye, enerji biyolojisine ve mekano-sinyal iletimine doğru derinleşti. Desminin mitokondri dağılımını ve solunum kapasitesini etkilediği; sarkomer-kostamer-nükleus hattında MAPK/PKC gibi yolların akışını biçimlendirdiği gösterildi. Kalpte ve iskelet kasında tekrarlayan gerilme-gevşeme döngülerine dayanıklılığı, desmin ağının yeniden düzenlenme kinetiği ve “çapraz konuşma”sıyla (aktin, mikrotübül ve nükleer lamina) sağlanıyordu. “Statik iskelet” tasviri yerini “dinamik bağ danteli”ne bıraktı.
2010’lardan itibaren desmin, post-translasyonel modifikasyonların (PTM) rafine bir “filament kodu”na sahip olduğu anlaşılan örnek proteinlerinden biri oldu. Baş-bölgesindeki serin/treonin fosforilasyonları, CaMKII-bağımlı işaretler ve O-GlcNAcylation arasındaki karşılıklı etkileşimlerin, filament montajı-dağılması ve sitoskeletal bölümlenme üzerinde zamansal-mekânsal kontrol kurduğu gösterildi. Bu çerçeve, kalp yetersizliği ve metabolik stres gibi durumlarda desminin neden bazen ağdan kopup agregasyona gittiğini, bazen de hücreyi kurtarıcı yanıtlarla stabil tuttuğunu açıklayan anahtar bir şemaya dönüştü.
Aynı dönemde moleküler ve klinik spektrum genişledi: “aynı aile içinde farklı kalp fenotipleri” üreten sıcak-bölgelerdeki (özellikle 1A alt-bölgesi) DES varyantları, iletim sistemi etkilenmesi ve ani kardiyak ölüm riskine uzanan çizgide haritalandı. Klinik elektrofizyolojiyle genetik arasındaki köprü güçlendi; fenotipik heterojenite, yalnızca mutasyonun konumuna değil, hücre iskeleti şaperonlarına, proteostaza ve PTM manzarasına da bağlandı. 2020’lerde, mozaik taşıyıcılığın bile desminopatilerde rol oynayabileceği gösterildi; bu da genetik danışmanlığın düz çizgisel öngörülerinin yeniden düşünülmesi anlamına geliyordu.
Tedavi ufku ise iki paralel patikadan ilerledi. Bir yanda, AAV-temelli gen aktarımı ve promotor hedeflemesi gibi stratejilerle “eksik desmini yerine koyma” veya hatalı ağı modüle etme girişimleri, fare modellerinde özellikle desminin tamamen yokluğunda kalpte anlamlı iyileşmeler gösterebildi. Diğer yanda, “fazla/yanlış desmin”in agregasyona eğilimli olduğu varyantlarda yalnızca daha çok desmin vermenin çözüm olmadığını; ağın doğru katlanması, doğru bağlanması ve doğru PTM imzasına kavuşması gerektiğini ortaya koyan çalışmalar, şaperon-modülasyon ve proteostaz hedeflerini gündeme taşıdı. Kısacası, desmin ailesinin klinikteki sorunları, “protein miktarı”ndan ziyade “protein kalitesi ve bağlamsal düzenlenmesi” meselesine dönüştü.
Patolojide desmin bugün de güçlü bir biyobelirteçtir. Rhabdomyosarkom ve leiomyosarkomda pozitif immünohistokimyasal boyanma, kas farklılaşmasının yol gösterici kanıtıdır; fakat güncel uygulama, desminin tek başına hüküm vermediğini bilir. İskelet kası farklılaşması için çekirdeksel myogenin ve MyoD1, düz kas için h-kaldesmon ve SMA ile oluşturulan panel, desminin “geniş kapı” duyarlılığını, “ince kapı” özgüllükle tamamlar. Bu bakımdan desmin, tanıda “ilk ipucu” olmayı günümüz moleküler patolojisiyle uyumlu biçimde sürdürür.
Bugün desmin araştırmaları, üç eksenin kesişiminde ilerliyor. Birincisi, PTM-kodunun çözülmesi: fosforilasyon-O-GlcNAcylation dengesinin filament mimarisine ve kalp ritim-kuvvet fizyolojisine etkileri, çok katmanlı omik yaklaşımlarla haritalanıyor. İkincisi, genetik-klinik spektrumun nüansları: mozaiklik, penetrans ve aile içi değişkenlik gibi konular yeni kohortlarda yeniden tanımlanıyor. Üçüncüsü, hedefli tedaviler: AAV temelli gen aktarımının doz-güvenlik sınırları, kardiyak özgüllüğü artıran promotorlar ve şaperon/otofaji yollarını “yeniden eğiten” farmakolojik stratejiler birlikte sınanıyor. Desminin adının ima ettiği “bağ”, bugün yalnızca z-disk ile sarkolemma arasında değil; temel bilim, patoloji, kardiyoloji ve gen tedavisi arasında da kuruluyor.
Kaynakça