
İçindekiler
Etimoloji ve tarihsel arka plan
“Pineal/pinealis” adı, bezin makroskopik koni/kozalak benzeri biçimine gönderme yapan Latince pinus (çam) ve pinea (çam kozalağı) köklerinden türemiştir; terim 16.–17. yüzyıl anatomi geleneğinde kalıcılaşmıştır. Erken modern dönemde epifiz, felsefe tarihinde René Descartes’ın zihin-beden etkileşiminin “tekil merkezi” olarak spekülatif biçimde anılmasıyla geniş bir kültürel görünürlük kazansa da, modern nörobilim bu yorumu desteklemez. 20. yüzyılın ortasında melatoninin izolasyonu ve kimyasal tanımlanması, epifizin sirkadiyen ve fotoperiyodik zamanlama ağlarındaki anahtar rolünü ortaya koyarak organın çağdaş biyomedikal önemini belirginleştirmiştir.
Topografik anatomi ve komşuluklar
Epifiz bezi, diensefalon çatısından köken alan epithalamusun bir parçasıdır. Üçüncü ventrikülün arka duvarına komşu pineal resese doğru uzanır; dorsalinde orta beynin dört tümsekten oluşan lamina quadrigemina (colliculus superior ve inferior çiftleri), ventralinde ise posterior kommissur bulunur. Orta hat yapısıdır; habenular kommissurla yakın ilişkisi, epithalamik ağ içinde konumunu pekiştirir. İnce bir bağ dokusu kapsülü ile sarılıdır ve parankim, stromal septalarla lobüllere ayrılır.
Damar yapısı ve drenaj
- Arteriyel beslenme: Başlıca posterior serebral arter dal sisteminden gelen medial ve lateral posterior koroidal arterler.
- Venöz drenaj: İç beyin venleri üzerinden vena cerebri magna (Galen veni) ve düz sinüse açılan hat.
İnnervasyon ve nöroanatomik bağlantılar
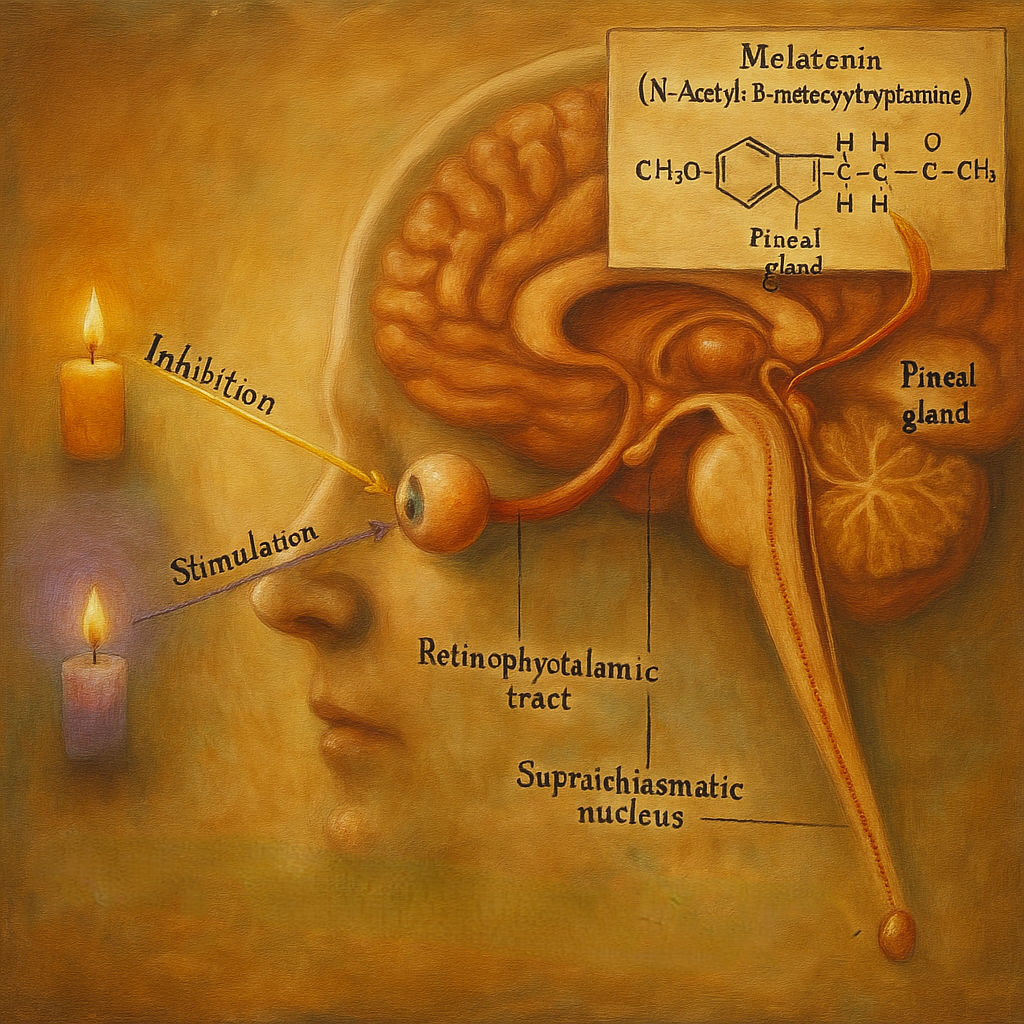
Epifizin ışığa duyarlı işlevi, doğrudan fototransdüksiyonla değil, retina kaynaklı sinyallerin hipotalamik saat devreleri üzerinden sempatik çıktı ile pineal parankime taşınmasıyla gerçekleşir:
- Retina melanopsinli ganglion hücreleri (ipRGC) →
- Suprakiazmatik çekirdek (SCN) →
- Paraventriküler çekirdek (PVN) →
- İnferior hipotalamik ve beyin sapı aracılığıyla inen otonom yol →
- Omuriliğin intermediolateral hücre kolonu (T1–T2) →
- Ganglion cervicale superius →
- Karotis pleksusu üzerinden epifize noradrenerjik lifler.
Parasempatik katkılar türler arası değişken ve ikincildir; memelide primer düzenleyici tonus sempatiktir.
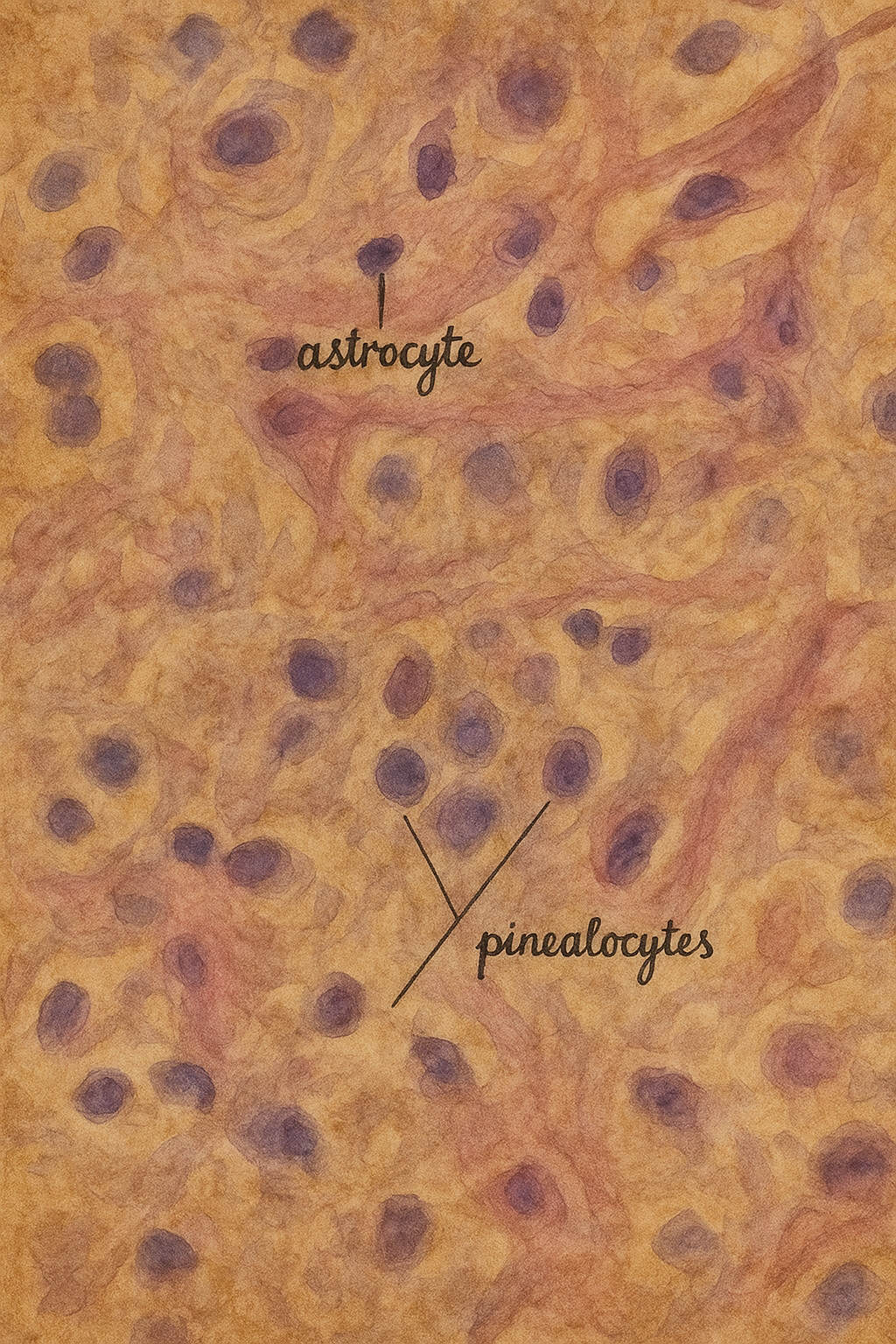
Histoloji ve hücresel organizasyon
- Pinealositler: Yuvarlak çekirdekli, zengin granüllü nöroendokrin hücrelerdir; uzantıları kapillerlere yaklaşır. Melatonin biyosentez enzimlerini (AANAT, ASMT vb.) yüksek düzeyde ifade ederler.
- İnterstisyel glia: Astrosit benzeri destek hücreleri, stromal mimarinin sürekliliğini sağlar.
- Stroma ve kapsül: Kollajen içeren bağ dokusu kapsülü ve ondan köken alan septalar, lobüler mimariyi belirler.
- Fizyolojik kalsifikasyonlar: Corpora arenacea (“beyin kumu”, akervulus) yaşla artma eğilimindedir. Klasik a.p. kranyografide orta hat referansı olarak parlak odak şeklinde görülebilir; güncel uygulamada BT/MR ile daha ayrıntılı değerlendirilir.
Fizyoloji ve biyokimya: melatonin sentezi ve sirkadiyen düzen
Işık-karanlık çevrimleri, epifize ulaşan sempatik tonusu değiştirerek melatonin üretimini zamana damgalar.
Işık sinyali ve hücre içi yolak
- Aydınlıkta: SCN aktivitesi sempatik çıkışı baskılar; pinealositlerde β-adrenerjik uyarı azalır, adenilat siklaz–cAMP–PKA ekseni düşer; melatonin sentezi azalır.
- Karanlıkta: Sempatik tonus artar; β1-adrenerjik reseptörler uyarılır; cAMP artışı AANAT enzimini transkripsiyonel ve posttranslasyonel düzeylerde etkinleştirir; sentez hızlanır.
Biyosentetik basamaklar
- Triptofan → 5-hidroksitriptofan → Serotonin (5-HT)
- Serotonin → N-asetilserotonin (hız kısıtlayıcı basamak; AANAT)
- N-asetilserotonin → Melatonin (ASMT/HIOMT)
Hedefler ve etkiler
- MT1/MT2 melatonin reseptörleri başlıca Gi/Go ile eşleşir; SCN nöral ateşleme paternlerini, vasküler tonusu, retina ve periferik saatlerin fazını modüle eder.
- Melatonin gece yükselen bir hormondur; DLMO (düşük ışık melatonin başlangıcı) bireysel sirkadiyen fazın biyobelirteci olarak kullanılabilir. Fotoperiyodik türlerde gün uzunluğuna göre mevsimsel (circannual) süreçlerin zamanlanmasına da aracılık eder.
Uyku-uyanıklık ve kronoterapi bağlamı
Melatonin, uyku başlangıcı kolaylaştırma ve sirkadiyen faz kaydırma amacıyla zaman/ışık hijyeniyle birlikte kullanılır. Etki, doz, uygulama saati ve aydınlatma koşulları arasındaki hassas ilişkiye bağımlıdır.
Anatomi notları: kapsül, komşuluk ve radyolojik korelat
Bağ dokusu kapsülü ve lobüler stroma, epifizin kontrast tutulumunu artırır; kan-beyin bariyerinin burada görece zayıf olması (fenestre kapillerler) MR’da belirgin kontrastlanma olarak yansır. Kalsifikasyonların sıklığı yaşla artar; BT’de hiperdenz, MR’da değişken sinyal paternleri görülür. Orta hat dışı sapmalar, kitle etkisi açısından ipucu verebilir.
Embriyoloji ve gelişim
Diensefalik çatının dorsal çıkıntısından 7.–8. gebelik haftalarında tomurcuklanır. Fetal ve erken çocukluk döneminde oransal olarak daha büyüktür; ergenlikten erişkinliğe parankimal küçülme ve kalsifikasyon artışı eğilimi gözlenir. Enzimatik saat genleri ve AANAT ritmi, doğum sonrası ışık çevresine uyumla olgunlaşır.
Evrimsel ve karşılaştırmalı biyoloji
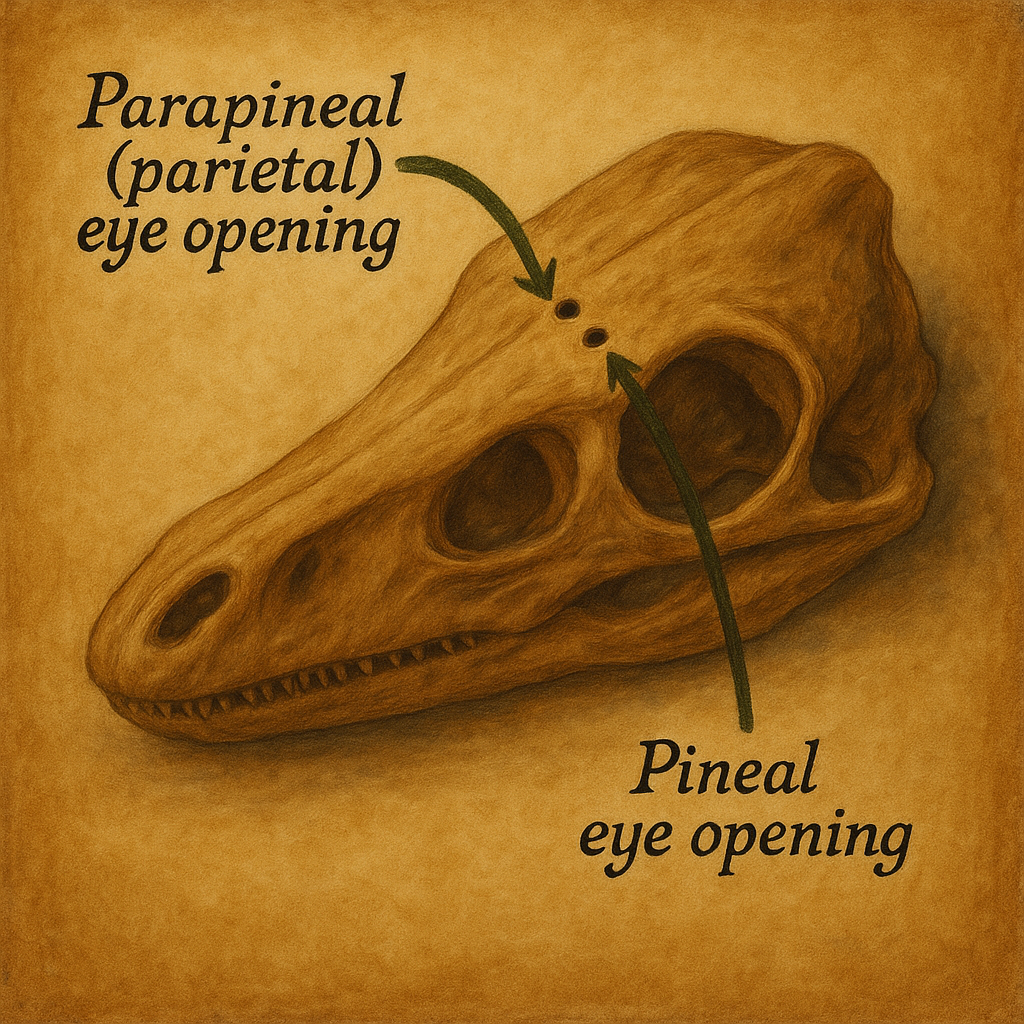
Alt omurgalılarda pineal kompleks sıklıkla doğrudan fotosensitiftir; bazı sürüngenlerde epifize komşu “paryetal göz” fotoreseptif yapı taşır. Bu türlerde ışık, kafatası üzerinden kısmen geçerek doğrudan pineal fototransdüksiyonu tetikleyebilir. Memelilerde ise kafatası/meninks ve beyin dokusunun ışığı engellemesi nedeniyle fotik bilgi retinohipotalamik yol üzerinden epifize dolaylı biçimde ulaşır. Bu evrimsel kayma, memelide retina/SCN merkezili sirkadiyen organizasyonun seçilimiyle uyumludur.
Klinik notlar ve patoloji
- Pineal kistler: Çoğu insidental ve asemptomatiktir; büyük lezyonlarda baş ağrısı, görsel yakınmalar veya aköz akım blokajına bağlı hidrosefali görülebilir.
- Pineal bölge tümörleri: Germ hücreli tümörler (ör. germinom) ve pineal parankimal tümörler (pineositom, PPID, pineoblastom) spektrumu; dorsal orta beyin basısına bağlı Parinaud sendromu (yukarı bakış felci, ışık-yakın ayrışması) tipiktir.
- Endokrin korelasyonlar: Epifize tarihsel olarak atfedilen “antigonadotropik/ergenlik karşıtı” işlev, modern insan verilerinde sınırlı ve bağlama duyarlıdır. Erken puberte epifiz fonksiyon bozukluklarında nadiren ve genellikle kitle etkisi ya da merkezi eksenlerin sekonder etkilenimiyle ortaya çıkar; melatoninin doğrudan ve genellenebilir bir “puberte baskılayıcı” olarak rutin kullanımı desteklenmez.
- Sirkadiyen bozukluklar: İleri/geri faz uyku-uyanıklık sendromları, körlükte serbest koşan ritimler, jet-lag ve vardiya tıbbında melatonin bir zamanlama işareti olarak değerlidir; ışık tedavisi ve davranışsal düzenlemelerle bütünleştirildiğinde etkilidir.
Görüntüleme ilkeleri
- BT: Kalsifikasyonları yüksek sensitiviteyle gösterir; kitlelerde heterojenlik ve komponent ayrımı.
- MR: T1/T2 sinyali yaş, kalsifikasyon ve kistik bileşime bağlıdır; gadolinyum sonrası belirgin kontrastlanma sık görülür.
- Radyografi: Güncel klinikte sınırlı olmakla birlikte, orta hat işareti olarak korpora arenacea geleneği tarihsel önemdedir.
Terminolojik düzeltmeler ve yazım notları
- “Sempatik boyun sınır ipliği” ifadesi, anatomik olarak truncus sympathicus ve özellikle ganglion cervicale superius kaynaklı noradrenerjik lifleri anlatır.
- “Ananasitler” yerine doğru terim pinealositlerdir.
- “Ergenlik karşıtı işlev” anlatımı tarihsel hipotezi yansıtır; güncel insan verisinde epifiz, pubertal gelişimi doğrudan engelleyen bir “fren” olmaktan ziyade, sirkadiyen/kronobiyolojik bağlamda modülatör bir etkene karşılık gelir.
Keşif
Antik başlangıç, bir “kapakçık” fikriyle başlar. İskenderiyeli Herophilos MÖ 4.–3. yüzyıllarda beyin karıncıkları kuramını geliştirirken çam kozalağını andıran bu küçük yapıyı, pneuma’nın (yaşamsal tin) akışını düzenleyen bir tür bekçi/vanaya benzetir; orta ventrikülden arka ventriküle geçişi “idare eden” bir kapı gibi. Galenos (MS 2. yüzyıl), hem diseksiyon titizliği hem de eleştirel yöntemiyle, iki önemli düzeltme getirir: Birincisi, epifizin beyin parenkiminden ayrı (diencephalon çatısında) ve üçüncü ventrikülün arkasında konumlandığını netleştirir; ikincisi, hareket etmeyen ve “dışarıda” duran bir yapının “kapak” olamayacağını gösterir. Bu yüzden ona, biçimini vurgulayan κωνάριον (konárion; Latince conarium) adını verir ve fonksiyon bulamayınca onu büyük beyin venlerine (sonradan “Vena Galeni”) destek veren bir “askı” gibi yorumlar. Bu Galenik temkin, Rönesans’a dek süren uzun bir “bilinmeyen organ” döneminin mihenk taşı olur.
Ortaçağ’da belleğin ve ruhun “yer” arayışı, metinlerde epifiz ile komşu yapılara (özellikle vermis cerebelli) dair bir isim karmaşası üretir. Qusṭā ibn Lūqā’dan Vincent of Beauvais’ye uzanan çizgide, bazen “pinea” adı solucanımsı vermis’e kaydırılır; “epifiz” ile “vermiform” terimleri yer değiştirir. Teolojik tartışmada ise Augustinus’un ruhun bedensel mekânı öğretisi, “ruhun beyin içindeki bir noktaya yerleşik olması” fikrini güçlendirir ve epifiz bu sembolik arayışa giderek daha sık dahil edilir.
Yeniçağ eşiğinde, Descartes bu küçük, eşlenmemiş bezde zihnin bedene bağlandığı “tekil menteşe”yi görür. “L’Homme” (özgün müsvedde 1630’lar; basım 1662/64) ile “Les Passions de l’âme”da (1649) epifizi “ruhun başlıca makamı ve düşüncelerimizin oluştuğu yer” diye niteler. Gerekçesi, anatominin simetri ilkesidir: Beynin çift taraflı yapılanışı içinde eşlenmemiş tek bir merkez, tüm duyumların birleştirildiği kavşak olabilir. Bu güçlü önerme, hem bilimsel hem kültürel tahayyülde üç yüzyıl yankılanır; fakat kısa süre sonra anatomi ve fizyolojinin yükselen eleştirel yöntemiyle tartışmaya açılır. Stensen (Niels Steno) ve Thomas Willis gibi çağdaşlar, “ruhun mekanı” tasarımını anatomik ve karşılaştırmalı gözlemlerle sorgular; epifizin hayvanlarda da “büyük ve güzel” olabildiğini, buna karşın “hayal gücü ve bellek” atıflarının ölçüsüz olduğunu hatırlatırlar.
19. yüzyıl, üçüncü gözün zoolojik keşfiyle hikâyeyi sarsıcı biçimde genişletir. Franz Leydig (1872), kertenkele beyninin ön–orta bölümünde “alın organı”nı tanımlar; Baldwin Spencer (1886) bazı sürüngenlerde epifizin bir sapla parietal foramen’e uzanıp “parietal göz” denen ışık alan bir yapıyla bağlantılı olduğunu gösterir. Nils Holmgren (1918), kurbağa ve köpekbalıklarında bu “parietal göz”ün retina koni hücrelerine benzeyen duyusal hücrelerden oluştuğunu, yani bezden çok fotoreseptif bir organ gibi davrandığını tarif eder. Böylece epifizin, alt omurgalılarda bir ışık-algılayıcı, üst omurgalılarda ise giderek “nöroendokrin” bir yapıya evrildiği fikri doğar. Richard Eakin’in “Üçüncü Göz” çalışmaları, bu karşılaştırmalı çizgiyi 20. yüzyılda popülerleştirir.
Klinik radyolojinin ve makroskopinin dili, aynı yüzyılda “beyin kumu” imgesini yerleştirir. Giovanni Battista Morgagni, epifizde yaşla artan kalsifiye taneleri “corpora arenacea” (beyin kumu) olarak adlandırır; bunların patolojik olmaktan çok fizyolojik çökeltiler olduğu, modern görüntülemede (direkt grafi, BT) bezin “işaret fişeği” gibi göründüğü anlaşılır. Yaşlanma, coğrafya ve bireysel farklılıklara göre değişen kalsifikasyon örüntüleri, melatonin senteziyle ilişkisi tartışmalı bir biyobelirteç haline gelir.
Fizyolojik devrim, 20. yüzyıl ortasında bir dermatoloji laboratuvarında başlar. 1958’de Aaron B. Lerner ve arkadaşları, sığır epifiz özlerinden kurbağa derisini hızla açığa çeviren bir indol türevi izole eder ve ona “melatonin” adını verir. Kısa sürede memelilerde bu molekülün gece–gündüz döngüsünü (sirkadiyen ritim) işaretleyen, çok-dokulu etkileri olan bir hormon olduğu gösterilir; epifiz böylece “son keşfedilen endokrin organ” payesine kavuşur. Julius Axelrod ve izleyicileri, melatoninin serotonin’den sentezinde hız kısıtlayıcı enzimin aralıklı salınım (gece artış) gösterdiğini, sempatik innervasyonla (üst servikal ganglion noradrenerjisi) geceleri AANAT/ASMT ekseninin “açıldığını” ortaya koyar. Bir tür “fotoneuroendokrin dönüştürücü” olarak epifiz, çevresel ışığı hormon diline çevirir.
Işığın bu dile nasıl “çevirildiği”, 1970’lerin beyin haritalarıyla keskinleşir. Retinadan hipotalamusa uzanan “retinohipotalamik trakt” tanımlanır; hipotalamusta suprakiazmatik çekirdek (SCN) memelilerin ana saat piyesi olarak yerini alır. SCN lezyonlarının ritimleri bozduğu, SCN’nin ışıkla ayarlandığı, SCN’den paraventriküler çekirdeğe, oradan omurilik aracı nöronlarına ve üst servikal gangliona, nihayet epifize uzanan çok basamaklı otonom yolun, melatonin sentezinin geceleyin “aç–kapa” anahtarı olduğu gösterilir. 1980’de Alfred Lewy ve ekibi, insanlarda geceleri karanlıkta yükselen melatoninin parlak ışıkla baskılandığını deneysel olarak ortaya koyar; insan sirkadiyen fizyolojisinde ışığın gücü ölçülür hale gelir.
Karşılaştırmalı biyoloji, kuş ve sürüngenlerde epifiz/“parietal göz” kompleksinin doğrudan ışığa duyarlı olabildiğini; memelilerde ise ışık bilgisinin retina–SCN–sempatik zincir üzerinden epifize ulaştığını gösterir. Modern nöroanatomi, epifizin esas girdisinin üst servikal ganglion kaynaklı sempatik lifler olduğunu; buna eşlik eden daha zayıf parasempatik liflerin (sfenopalatin/otik ganglion) peptiderjik düzenleyiciler taşıyabildiğini haritalar. Bu nöral mimari, beta-adrenerjik blokajın melatonin gece sivrisini bastırabilmesi, servikal sempatik lezyonlarda ritmin zayıflaması, tek taraflı innervasyon deneylerinde iç karotis sinirinin özel rolü gibi bulgularla desteklenir.
Biyokimyasal zaman işaretinin klinik izdüşümleri genişler. Mevsimsel üreme fiziolojisinde melatonin sinyali, gün uzunluğunu gonad eksenine “çevirir”; insanda ise gecikmiş uyku fazından kör bireylerde sirkadiyen serbest koşuya, vardiya uyku bozukluklarından jet-lag’a dek pek çok durumda zamanlayıcı ya da “kronobiyotik” bir araç olarak kullanılır. Nöroimmünoendokrinoloji, melatonine membran reseptörleri (MT1/MT2) yanında çekirdek/mitokondri düzeyinde bağlanma yerleri atfederek antioksidan, immünmodülatör, nöroprotektif işlevleri tartışır. Yine de insan yaşlanmasında epifiz kalsifikasyonu–melatonin üretimi–uyku kalitesi arasındaki ilişkiler heterojen ve nüanslı bulunur; ülkeler arası varyasyon ve bireysel kronotip farklılıkları, tek değişkenli açıklamaları sınırlar.
Bu uzun hikâyeden geriye, Descartes’ın sezgisel simetrisi ile Galenos’un anatomik ihtiyatı arasında sürüp giden bir diyalog kalır: Epifiz, alt omurgalılarda bir “ışık-alma” harkuladeliliğinden, memelilerde ışığı otonom sinyale, o sinyali gece hormonuna çeviren bir transdüksiyon düğümüne evrilmiştir. Ruhun tahtı arayışı ise, yerini fotonların sinirsel koduna, o kodun da endokrin zamana çevrildiği, çok katmanlı bir biyolojiye bırakmıştır.
İleri Okuma
- Erlich, S.S. (1985). The pineal gland: anatomy, physiology, and clinical significance. Journal of Neurosurgery, 63(3), 321–341. https://doi.org/10.3171/jns.1985.63.3.0321 (thejns.org)
- Kvetnoy, I. et al. (2022). Melatonin as the Cornerstone of Neuroimmunoendocrinology. International Journal of Molecular Sciences, 23(3), 1–39. https://doi.org/10.3390/ijms23031092 (PMC)
- Lewy, A.J.; Wehr, T.A.; Goodwin, F.K.; Newsome, D.A.; Markey, S.P. (1980). Light suppresses melatonin secretion in humans. Science, 210(4475), 1267–1269. https://doi.org/10.1126/science.7434030 (cet.org)
- Moore, R.Y.; Lenn, N.J. (1972). A retinohypothalamic projection in the rat. Journal of Comparative Neurology, 146(1), 1–14. https://doi.org/10.1002/cne.901460102; Stephan, F.K.; Zucker, I. (1972). Circadian rhythms in the rat: deep lesions of the suprachiasmatic nuclei. Science, 178(4058), 1325–1327. (PMC)
- Reiter, R.J. (2010). Melatonin: A multitasking molecule. Advances in Experimental Medicine and Biology, 695, 1–15. https://doi.org/10.1007/978-1-4419-6247-6_1 (sciencedirect.com)
- Tan, D.-X.; Hardeland, R.; Manchester, L.C.; et al. (2018). Pineal calcification, melatonin production, aging, associated health consequences and rejuvenation of the pineal gland. Aging and Disease, 9(5), 1–31. https://doi.org/10.14336/AD.2017.1215 (PMC)
- Møller, M.; Bock, E. (2002). The anatomy and innervation of the mammalian pineal gland. Microscopy Research and Technique, 59(6), 409–426. https://doi.org/10.1002/jemt.10217 (PubMed)
- Bowers, C.W.; Zigmond, R.E. (1984). The number and distribution of sympathetic neurons that innervate the rat pineal gland. Cell and Tissue Research, 236, 127–134. https://doi.org/10.1007/BF00217101 (PubMed)
- Whitehead, M.T.; Oh, C.C.; Choudhri, A.F. (2015). Physiologic Pineal Region, Choroid Plexus, and Dural Calcifications in Children on CT. Radiology Case Reports, 10(3), 1–5. https://doi.org/10.2484/rcr.v10i3.1030 (PMC)
- Eakin, R.M. (1973). The Third Eye. University of California Press, Berkeley. ISBN 9780520021737. (Bk. ayrıca Leydig, 1872; Spencer, 1886; Holmgren, 1918 göz/epifiz literatürü derlemesi.) (Wikipedia)
- Stanford Encyclopedia of Philosophy (2020, Fall). Descartes and the Pineal Gland. In: Zalta, E.N. (Ed.), SEP. (plato.stanford.edu)
- “History of the pineal gland.” (2025). Wikipedia (derleme ve kaynakça taraması için). (Wikipedia)