Sinonim: Vacuole.
- Latincedeki vacuus (“boş”)’un küçültme eki alan biçimi vacuola‘ya, Fransızcada vacuole denir.
- Su gibi anorganik ve organik molekülleri içeren etrafı kapalı organele denir.
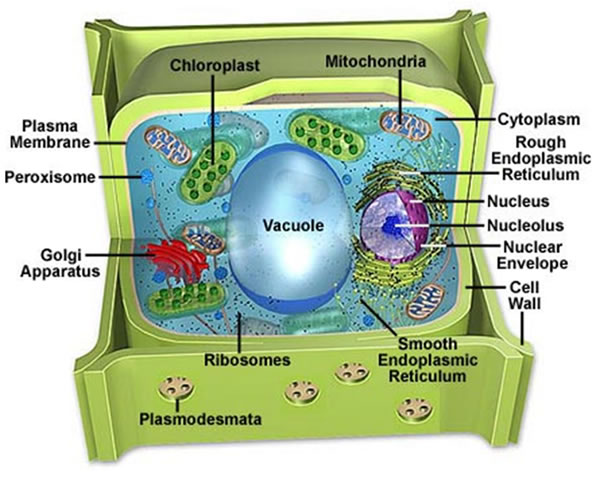
Tıp terimleri sözlüğü
Sinonim: Vacuole.

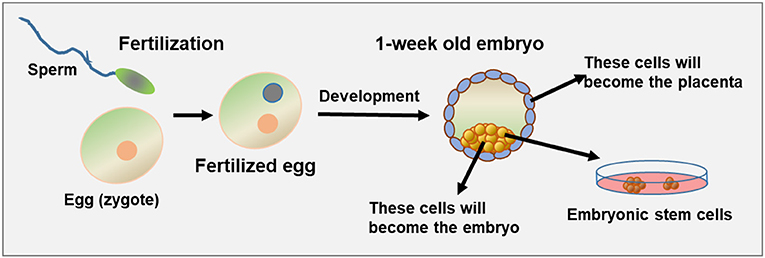
İnsanlar, trilyonlarca yüksek düzeyde özelleşmiş hücreden oluşan ökaryotik çok hücreli organizmalardır. Bu hücrelerin tümü, tek bir döllenmiş yumurta hücresinden (zigot) bölünerek üretilir. Her organın farklı temsilcileri vardır, örneğin kas hücreleri, kalp kası hücreleri, karaciğer hücreleri, bağırsak hücreleri ve sinir hücreleri. Vücut, 200’den fazla farklı hücre tipine ev sahipliği yapmaktadır.
Kök hücreler, kendilerini yenileyen ve bu özelleşmiş hücrelerin öncüsü olarak gelişen hücrelerdir. Vücut hücrelerinin aksine, herhangi bir sayıda bölünebilirler ve potansiyel olarak ölümsüzdürler.
Bir yandan organizmayı oluştururlar ve sürekli yenilenme için ona hizmet ederler. Örneğin cilt yaklaşık ayda bir yenilenir.
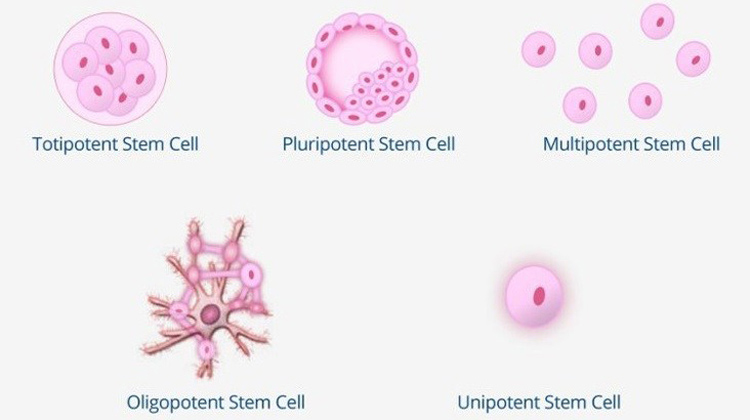
Çok hücreli bir organizmanın ömrü boyunca, yetişkin kök hücreleri denilen gösterilebilir. Rrganlarda, kemik iliğinde, aynı zamanda göbek kordonunda da bulabilirsiniz. Embriyonal kök hücreler kadar ücretsiz olarak uzmanlaşamazsınız ve kendilerini bu dokunun hücrelerine ayırarak (onarım) kendilerini ayırt ederek altta yatan dokuyu değiştirme işlevine sahip olabilirsiniz. Bununla, bölünme yeteneğinizi kaybedebilir, ancak dokunun belirli görevlerini yerine getirebilirsiniz. Yetişkin kök hücreler genellikle artık kendilerini herhangi bir dokuda ayırt etme yeteneğine sahip değildir, ancak isteğe bağlı olarak sadece bir dokunun farklı hücre tiplerinde. Bu nedenle çok parçalı olarak adlandırılırlar.

Embriyonal kök hücreler iki türe ayrılabilir:
Farklı kök hücre türleri vardır.
Sözde uyarılmış pluripotent kök hücreler (iPS hücreleri, iPSC), örneğin bir deri hücresi gibi somatik hücrelerden elde edilebilir. Sözde ‘yeniden programlanırlar’ ve embriyonik kök hücrelerle karşılaştırılabilir özelliklere sahiptirler. Avantajı, kabulü artıran hiçbir embriyoya gerek olmamasıdır.
Somatik hücre nükleer transferinde (SCNT), somatik bir hücrenin çekirdeği, çekirdeksiz bir hücreye aktarılır. Örneğin, embriyonik kök hücre kaynağı olarak hizmet eden bir blastosistin geliştiği yumurta hücrelerinde.
Bir başka kök hücre kaynağı da yenidoğanın göbek bağıdır.
Hücreler yaşamın temel yapı taşları olduğundan, kök hücreler tıpta büyük bir tedavi potansiyeline sahiptir. Örneğin, insülin üreten beta hücreleri nakledilebilir ve böylece tip 1 diyabeti tedavi edebilir. Veya felç, sinir hücrelerinin yenilenmesiyle tersine çevrilebilir.
Örnekler:

Kök hücre kullanmanın potansiyel tehlikeleri arasında kanser ve teratom gelişimi yer alır. Vücuda yabancı kök hücreler kullanıldığında, bağışıklık sisteminin aracılık ettiği reddedilme reaksiyonları riski vardır. Ancak otolog hücreler söz konusu olduğunda bu tehlike mevcut değildir.
Unutulmamalıdır ki, kök hücre konusunda yetkililer tarafından onaylanmayan şaibeli teklifler vardır. Bunlar hesaplanamaz riskler barındırdıkları için kullanılmamalıdır.
Embriyonik kök hücrelerin araştırma ve tıpta kullanımı etik olarak tartışmalıdır, çünkü onları elde etmek için sözde yıkıcı embriyo araştırması, yani embriyonun öldürülmesi gerekir. Bununla birlikte, kök hücre araştırmalarını tabu haline getirmek, terapötik ilerlemeyi engelleyebileceği kadar tartışmalıdır.

Ayrıca bitkilerin kök hücreleri var. Bunlar, filizin üstünde, sözde apikal stres olarak ve köklerdeki kök zirvelerinde bulunur. Bununla birlikte, hayvan ve insan hücrelerinin aksine, bitkiler hemen hemen tüm hücreler, tam bir organizmayı yenileme yeteneğine sahiptir.

Sinonim: Pilz.
Latincede fungus (tekil; mantar), fungi (çoğul; mantarlar). Çok hücreli ve tek hücreli olabilen ökaryotik canlıları kapsayan canlılar alemidir.

Sinonim: Gewebe, tissue
Hayvan, bitkilerde hücrelerin belirli bir işlevi gerçekleştirmek için bir araya gelmesidir.

Kulüp hücresi, memelilerin alt solunum yollarında—özellikle terminal ve respiratuvar bronşiollerde—yerleşik, kirpiksiz, kubbemsi apikal yüzeye sahip bir sekretuvar epitel hücresidir. İngilizce literatürde “club cell” olarak anılır; tarihsel eponim “Clara cell” günümüzde etik nedenlerle terk edilmiştir. Latince karşılığı Exocrinocytus bronchiolaris dizgesel adlandırmayı vurgular ve hücrenin bronşiolar epiteldeki ekzokrin işlevini öne çıkarır.
Hücre ilk kez 1937’de Alman anatomist Max Clara tarafından tanımlanmıştır. Clara’nın Nazi Partisi üyeliği ve etik dışı örnek kullanımlarına dair bulgular, modern solunum biyolojisi topluluğunda eponimin sürdürülmesini uygun görmeyen bir konsensüs doğurmuştur. Bu nedenle 21. yüzyılda büyük dergiler ve alan kılavuzları, “Clara hücresi” yerine ideolojik çağrışımı olmayan “kulüp (club) hücresi” terimini benimsemiştir.
Kulüp hücreleri, trakea ve büyük bronşların değil, intrapulmoner küçük havayollarının belirgin elemanlarıdır. En yoğun olarak:
Işık mikroskopisinde apikal yüzeyi hafifçe bombeleşmiş, kirpiksiz, sitoplazması eozinofilik bir hücre görünümü verir. Elektron mikroskopisinde:
Kulüp hücrelerinin ayırt edici belirteci SCGB1A1 (eski adlarla CC10/CC16, uteroglobin) proteinidir; immünohistokimyada güçlü sitoplazmik boyanma verir. Kirpikli hücre belirteci FOXJ1 ifadesi yoktur. Türler arasında değişmekle birlikte, CYP ailesinden izoenzimler (örn. insanlarda CYP2F1/2B6 ailesi; kemirgenlerde CYP2F2) kulüp hücrelerinin ksenobiyotik metabolik kapasitesinin temelini oluşturur. MUC5AC gibi jel-forming müsinler esasen goblet hücrelerine aittir; kulüp hücresinde bazal/iz düzeyde görülür.
Embriyogenezde proksimo-distal hava yolu paternlenmesinin bir parçası olarak, endoderm kökenli hava yolu progenitörlerinden farklılaşırlar. Erişkin akciğerde kulüp hücresi, bronşiolar epitelin uzun ömürlü progenitörü işlevini görür:
Kulüp hücreleri, SCGB1A1/CC16 başta olmak üzere anti-inflamatuvar, antiproteaz ve yüzey düzenleyici proteinler salgılar. Alveoler tip II hücreleri surfaktantın ana kaynağı olsa da, kulüp hücreleri bronşiolar lümende fosfolipid ve protein bileşenlerle yüzey gerilimi ve sıvı tabakasının viskoelastik özelliklerine katkıda bulunur.
Gelişkin sitokrom P450 donanımı sayesinde inhalasyonla alınan hidrofobik kimyasalları metabolize eder. Bu çift taraflı bir kılıçtır: Bazı toksinler bizzat kulüp hücresinde biyolojik olarak etkin ara ürünlere dönüştürülerek selektif kulüp hücresi nekrozuna yol açabilir (kemirgenlerde naftalen modeli klasik örnektir).
Hücre–hücre bağlantıları ve bazal hücrelerle kurduğu etkileşimler aracılığıyla bronşiolar epitelin geçirgenliğini düzenler; mitotik aktivasyonla hasar sonrası epitel yenilenmesine katılır. Niş faktörleri (ör. Wnt, Notch, Hippo-YAP sinyallemesi) farklılaşma yönünü belirler.
SCGB1A1/CC16 ve ilgili sekretuvar ürünler, nötrofil elastazı ve fosfolipaz A2 gibi enzimleri inhibe ederek inflamasyonu yatıştırır; epitel–immün hücre arayüzünde sitokin akışını dengeler. Bu özellik, astım ve viral bronşiolit gibi durumlarda klinik yansımalar taşır.
Kemirgenlerde kulüp hücreleri çok bol ve naftalene aşırı duyarlıdır; insanlarda benzeri metabolik yollar mevcuttur ancak izoform dağılımı ve hassasiyet farklıdır. İnsan küçük havayollarında bazal hücreler de anlamlı bir progenitör havuz oluşturur; bu, onarım dinamiklerini türler arasında değişken kılar.
Serum ve balgamda ölçülebilen CC16, bronşiolar epitel bütünlüğünün duyarlı bir göstergesi olarak çalışır.
Farelerde naftalen ve belirli klorlu hidrokarbonlar kulüp hücrelerini seçici olarak hedefler. Bu modeller, küçük havayolu onarımını, niş sinyallerini ve progenitör davranışlarını çözümlemede standarttır. İnsanlarda benzer biyokimyasal eksenler olsa da doz–yanıt ve enzim izoform dağılımları farklılık gösterir.
Kulüp hücresi—latinceleştirilmiş adlandırmasıyla Exocrinocytus bronchiolaris—küçük havayolu epiteline özgü kirpiksiz, kubbemsi, sekretuvar bir epitel hücresidir. Hikâyesi, klasik ışık mikroskobisinin sınırlarıyla başlayan morfolojik bir merakın, elektron mikroskobisi çağında ayırt edici ultrastrüktüre dönüşmesi, daha sonra biyokimya ve toksikoloji ile işlev kazanması ve nihayetinde modern soy-izleme, organoid ve tek-hücreli transkriptomiklerle yeniden yazılmasına uzanır.
1930’ların sonunda Max Clara, insan ve tavşan bronşiyollerinde kirpiksiz, apikal yüzeyi kubbemsi, sitoplazması granüllü hücreleri sistematik biçimde tanımladı; apikaldeki granüller ve zengin endoplazmik retikulum ağını vurgulayan bu tanımlama hızla benimsenerek yapıtaşa adını verdi. Sonraki on yıllarda, “Clara hücresi” eponimi anatomi ve patoloji atlaslarının standart terminolojisine dönüştü; küçük havayolu epiteli artık kirpikli, goblet ve “Clara” hücresinden oluşan üçlü bir şema üzerinden anlatılıyordu.
İkinci Dünya Savaşı sonrası elektron mikroskobisinin biyolojiye girmesi, kulüp hücresinin “ne gördüğümüzü” netleştirdi: apikalde elektron-yoğun sekretuvar granüller, geniş lamelli düz endoplazmik retikulum, sınırlı mikrovilluslar ve komşu hücrelerle sıkı bağlantı kompleksleri. Bu ultrastrüktürel imza, kulüp hücresini yalnızca “kirpiksiz” olmakla değil, sekresyon ve metabolik işlevlere adanmış bir organel mimarisiyle ayırt etmeyi mümkün kıldı. Aynı dönemde bronşiol-alveol geçiş bölgesinde (BADJ) bu hücrelerin sayıca arttığı ve goblet hücrelerinin tersine proksimo-distal eksende belirginleştiği gösterildi; bu topografya, fonksiyonel rolleri hakkında ilk ipuçlarını verdi.
1960’lardan itibaren iki paralel hat belirginleşti. İlki, kulüp hücresinin karakteristik sekretuvar ürünlerine odaklandı: bugün SCGB1A1 olarak adlandırılan ve klinikte CC16/CC10 biçiminde ölçülen küçük, anti-inflamatuvar bir protein bu hücrenin ayırt edici imzası oldu. İkincisi, düz endoplazmik retikulumun barındırdığı sitokrom P450 enzim kümelerinin uçucu organik bileşikler ve çevresel toksinlerin biyotransformasyonunda başrol oynadığı anlaşıldı. Bu bilgi, deneysel toksikolojide bir kavşak açtı: bazı hidrokarbonların (örnek model: naftalen) önce kulüp hücresinde reaktif metabolitlere çevrilip selektif nekroz oluşturması, bu hücreyi küçük havayolu hasarı ve onarımının “doğal işaretleyicisi”ne dönüştürdü. Böylece kulüp hücresi, hem detoks yapan “bekçi”, hem de kimi ajanlara karşı “ilk hedef” sıfatlarını birlikte taşıyan bir paradoksun merkezi haline geldi.
1990’lar ve 2000’lerin başı, akciğer gelişim biyolojisinin moleküler araçlarla hızlandığı dönemdir. SCGB1A1 promotoruna dayalı genetik işaretleme ve Cre/loxP tabanlı soy-izleme yaklaşımları, erişkin bronşioler epitelde kulüp hücrelerinin uzun ömürlü progenitör işlevi gördüğünü ve hasar sonrası kirpikli hücrelere sınırlı dönüşüm kapasitesi taşıdığını gösterdi. Bronşiol-alveol kavşağında, tip II pnömositlerle komşu “niş” alt kümelerinin onarımda göreli olarak daha baskın rol üstlendiği ortaya kondu. Bu bulgular, küçük havayolunu statik bir mozaik değil, yarı-stabil bir doku devri-daim sistemi olarak düşünmeye zorladı.
Sekretuvar-kirpikli kader ikiliğini belirleyen sinyal yolları giderek netleşti. Notch aktivitesi sekretuvar kimliği (kulüp fenotipini), Notch baskılanması ise kirpikli farklılaşmayı destekleyen bir anahtar gibi davrandı. Wnt/β-katenin ve Hippo-YAP/TAZ eksenleri doku gerilimi, yara onarımı ve hacim koruma süreçlerinde kulüp hücresinin çoğalım/diferansiyasyon kararlarını ayarlayan bağlamsal düğümler olarak öne çıktı. Bu çerçeve, inhalasyon hasarı sonrasında neden bazen sekretuvar havuzun hızla genişlediğini, bazense kirpikli yola akışın arttığını açıklamakta kullanıldı.
Kulüp hücresinin sekresyon ürünü CC16, hem serumda hem balgamda ölçülebilirliği sayesinde bronşioler epitel bütünlüğünün bir “uzaktan göstergesi” haline geldi. Sigara dumanı maruziyeti, küçük havayolu daralması ve KOAH spektrumunda düşük CC16 düzeyleri sıklıkla saptandı; akut akciğer hasarında ise bariyer sızıntısına bağlı geçici yükselmeler gözlendi. Bu biyobelirteçlik, işyeri maruziyetlerinden viral bronşiolite kadar geniş bir yelpazede epidemiyolojik ve klinik araştırmalara yakıt sağladı. Aynı zamanda insan akciğerinde bazal hücrelerin de anlamlı bir progenitör havuz oluşturduğu fark edilerek, “insan-kemirgen farkları” başlığı altında kulüp hücresinin göreli payının doku ve tür bağlamına duyarlı olduğu kabul edildi.
Max Clara’nın biyografisine ilişkin etik sorunlar ve dönemin insan dokusu kullanımına dair bulgular, 21. yüzyılda alanın önde gelen dergi ve derneklerinde eponimin terk edilmesi yönünde güçlü bir norm doğurdu. Böylece ideolojik çağrışımlardan arındırılmış “kulüp (club) hücresi” adı hızla yerleşti; terminoloji, hücrenin şekilsel benzetmesinden (apikal kubbelenme/kulüp başı imgesi) ve sekretuvar doğasından esinlenen, betimleyici ve işlevsel bir anlatıma döndü.
Son on yılda tek-hücreli RNA dizileme ve mekânsal transkriptomik, kulüp hücresini homojen bir popülasyon olmaktan çıkarıp fonksiyonel alt kümelere ayırdı: metabolik olarak yüksek P450 yüklü alt tipler; immün-modülatif imza baskın, SCGB-zengin alt tipler; onarıma meyilli, geçici olarak proliferatif alt tipler. Bronşiol-alveol kavşağı, nöroepitelyal cisimcik komşuluğu ve küçük havayolu dallanma noktaları gibi mikronişler, bu alt tiplerin mekânsal dağılımını belirleyen haritalar olarak çizildi. İnsan dokusunda bazal hücrelerin eşlik eden progenitör rolü ve kronik maruziyetlerde goblet metaplazisi ile kulüp havuzundaki görece azalma, hastalık fenotipleriyle ilişkilendirildi.
Hava-sıvı ara yüzeyli (ALI) primer kültürler ve organoid sistemler, insan kökenli kulüp fenotipinin in-vitro yeniden kurulmasına ve ilaç/toksisite taramalarına olanak verdi. CRISPR tabanlı soy-izleme ve reportör sistemleri, hasar-onarım döngülerinde kulüp hücresinin kaderini tek-klon çözünürlüğünde izlemeyi mümkün kıldı. Bu teknikler, örneğin aerosolize tedavilerin küçük havayolu hedeflemesi, mukosiliyer arayüzün viskoelastik ayarı ve epitel-immün etkileşimlerin incelenmesi gibi çeviri sorularına doğrudan hizmet etmeye başladı.
Bugün araştırmalar üç ana eksende yoğunlaşıyor.
Kısacası, Kölliker’in ışık mikroskobu altında “farklı” diye işaretlediği hücresel siluet, Clara’nın sistematik tanımıyla isim kazanmış; elektron mikroskobunda ultrastrüktür, toksikolojide reaktivite, biyokimyada CC16 imzası ve gelişim biyolojisinde progenitörlük rolleri ile işlevlenmiş; tek-hücreli ve mekânsal teknolojiler çağında ise alt-tip, niş ve kader haritalarıyla zenginleşmiştir. Bugün “kulüp hücresi” adıyla anılan bu yapıtaşı, küçük havayolunun çevresel maruziyetlere verdiği yanıtın, bariyer bütünlüğünün ve onarım dinamiklerinin merkezindeki aktörlerden biridir.
Sinonim: gland, Drüse
Vücut yüzeyine, boşluğuna veya damar ağının içine madde sentezleyip salgılayan organdır.
1)Ekzokrin salgı bezleri; ter bezleri, yağ bezleri, süt bezleri, sindirim bezleri, mukus bezleri v.b.
2)Endokrin salgı bezleri
1)Merokrin bezi (Sin: Merocrine (Eccrine) gland, ekkrine Drüse): Salgılama sırasında bu bezdeki hücreler kendilerinden hiç kayıp vermeden veya az kayıp vererek salgı oluştururlar. Ekrin bezleri özellikle ter bezlerini ifade etmek için kullanılır.(Bkz; Ek–krin) (Bkz; Mero–krin)
2)Apokrin bezi (Sin: Apocrine gland, Apokrine Drüse): Salgılama sırasında bu bezdeki hücrelerin bir kısmı salgıya dönüşür. (Bkz; Apo–krin)
Örneğin: Süt bezi.
3)Holokrin bezi (Sin: Holocrine gland, holokrine Drüse): Salgılama sırasında bu bezdeki hücrelerin tamamı salgıya dönüşür. (Bkz; Holo–krin)



İnsanlar neredeyse çırılçıplak durumdaki derileriyle, primatlar arasında benzersizdir. Sülalemizin tüm diğer üyeleri, çoğu başka memeli türünde olduğu gibi kalın bir kürke sahiptir. Biz insanların da kafasında ve birkaç başka yerinde daha kıl olduğu doğru. Ama yine de akrabalarımıza kıyasla en kıllı insan bile kel sayılabilir.
Peki nasıl bu kadar kılsızlaşabildik? Bilimciler yüzyıllar boyunca bu soruya kafa yordu. Yanıt bulmak ise kolay değildi. İnsan evrimindeki en önemli geçişler (örneğin dik yürüyüşün ortaya çıkışı gibi), atalarımızın fosillerinde doğrudan kayıt altına alınmış durumdaydı. Fakat kalıntıların hiçbirinde insan derisine ilişkin kesin izler korunamamıştı.
Neyse ki son yıllarda araştırmacılar, kıllarımızı yitirmemize ilişkin birtakım dolaylı ipuçlarının fosil kayıtlarında mevcut olduğunun farkına vardı. Bu ipuçlarının ve geçtiğimiz on yılda genomik ile fizyoloji sayesinde kazanılan anlayışın ışığında, antropolog Nina G. Jablonski ve çalışma arkadaşları insanların ne zaman ve neden kürklerinden kurtulduklarını ikna edici biçimde açıklamayı başardı. Görünüşümüzün garipliklerinden birini açıklamasının yanı sıra, bu senaryoya göre kılsız derinin kendisi de diğer karakteristik insan özelliklerinin evriminde rol oynamış. Buna beyin büyüklüğümüz ve konuşma diline bağımlılığımız da dahil.
Atalarımızın vücut kıllarını neden yitirdiklerini anlamak için öncelikle diğer türlerin neden kürklerinin olduğunu düşünmemiz gerek. Kıllar, memelilere özgü bir vücut örtüsüdür. Bu canlı grubunun tanımlayıcı özelliklerinden birinin kürk olduğuna kuşku yoktur: Neredeyse tüm memeliler hiç değilse bir miktar kıla sahiptir; çoğunun da yoğun bir kürkü olur. Kıllar sıyrılmaya, ıslanmaya, güneşin zararlı ışınlarına ve potansiyeler tehlikeleri olan parazit ya da mikroplara karşı yalıtım ve koruma sağlar. Ayrıca yırtıcıların kafasını karıştıracak bir kamuflaj görevi görür. Ayırt edici desenleri sayesinde de aynı türün üyelerinin birbirlerini tanımalarını kolaylaştırır. Dahası, memeliler kürklerini saldırganlık ya da huzursuzluk ifade etmek için de kullanırlar: Örneğin bir köpek boyun kıllarını kabarttığında, karşısındakine açık bir “uzak dur” sinyali vermiş olur.
Her ne kadar kürk böylesine önemli amaçlara hizmet etse de, bazı memeli soylarında son derece seyrek ve işlevsiz kıllar bulunur. Bu türlerin büyük bölümü yeraltında veya suda yaşayan canlılardır. Yeraltı memelilerinde, örneğin “çıplak kör fare” adı verilen kemirgenlerde, kılsızlığın evrimi yeraltında büyük koloniler halinde yaşamaya yanıt olarak gelişmiştir. Bu yaşam biçiminde kürk fazlalık duruma gelir; çünkü hayvanlar zaten karanlıkta birbirlerini görmezler ve sosyallik de ısınmak için bir araya toplanmalarından ibarettir. Kıyıya hiç çıkmayan deniz memelilerinde, örneğin balinalarda, kılsızlık deri yüzeyindeki sürtünmeyi azaltarak, uzun mesafelere yüzmeyi ve dalmayı kolaylaştırır. Dış yalıtımın yokluğunu telafi etmek için bu hayvanların derilerinin altında yağ katmanı olur. Yarı sucul memelilerde, örneğin su samurlarında ise genellikle tersine yoğun ve su geçirmez bir kürk olur. Bu kürk kaldırma kuvveti sağlamak için havayı içine hapsederek, su üzerinde kalmayı kolaylaştırır. Tabi aynı zamanda karadayken deriyi de korur.
Karasal memelilerin en büyükleri de -yani filler, gergedanlar ve su aygırları- kılsız deriler geliştirmiştir; çünkü sürekli olarak aşırı ısınma riski ile karşı karşıyadırlar. Bir hayvan ne kadar büyük olursa, toplam kütlesine oranla o kadar az yüzey alanına sahip olur. Bu nedenle vücudunu fazla ısıdan kurtarması güçleşir. Öte yandan fareler ve diğer küçük memelilerin yüzey/hacim oranı yüksek olduğundan, çoğu zaman yeterli ısıyı koruma mücadelesi verirler. Yaklaşık 2 milyon yıl öncesinden 10.000 yıl öncesine kadarki zamanı kapsayan Pleistosen Devri sırasında, mamutlar ile modern fillerin diğer akrabalarının ve ayrıca gergedanların kürkleri vardı; çünkü o zamanlar soğuk bir ortamda yaşıyorlardı. Dış yalıtım vücut ısılarını korurken, besin gereksinimini de azaltıyordu. Ancak günümüzde tüm mega otçullar çok sıcak koşullarda yaşıyorlar; dolayısıyla kürk onlar için ölümcül olurdu.
İnsanın çıplaklığı ise ne yeraltında ne de suda yaşama uyumlanmanın bir sonucu değildir. Bir zamanların gözde “sucul maymun” varsayımı da artık geçerliliğini yitirdi. Çok büyük boyutlu olmaktan ileri gelen bir kürksüzlük olmadığı da kesin. Bizim kürksüz bedenimiz, üstün terleme yeteneğimiz göz önüne alınırsa, serin kalabilmekle ilgili görünüyor.

Serin kalmak, yalnızca iri yapılı olanlar için değil pek çok memeli için büyük bir problemdir, özellikle de sıcak bölgelerde yaşayan ve uzun yürüme ya da koşma mesafelerinde aşırı miktarda ısı üretilir. Bu hayvanlar vücut sıcaklıklarının dikkatli bir biçimde düzenlemelidir, çünkü dokuları ve organları, özellikle de beyinleri, aşırı ısıdan dolayı hasar görebilir.
Memeliler aşırı ısınmaya karşı çeşitli taktikler geliştirmiştir. Örneğin, köpekler hızlı bir biçimde soluk alıp verirler, pek çok kedi türü genellikle daha serin olan akşam saatlerinde aktiftir ya da antilopların çoğunluğu burunlarından nefes alarak burunlarında bulunan küçük damarlar aracılığıyla damarlarındaki kanı soğutur. Fakat insanların da içerisinde yer aldığı primatlarda ise terleme, birincil stratejidir. Terleme, deri yoluyla atılan suyun vücut dışına çıktığında deriden aldığı ısı ile buharlaşması sonucu deriye serinlik kazandırır. Bütün bu vücut soğutma mekanizması, aynı prensibe (buharlaşma ile serinleme) göre çalışır, beyin ve diğer organların aşırı ısınmadan dolayı hasar görmesini engellemede ustaca geliştirilmiş bir yöntemdir.

Fakat bütün terlemeler aynı değildir. Memeli derisi, üç tip bez bulundurur; sebasöz, apokrin ve ekrin. Bu bezler birlikte çalışarak ter üretirler. Birçok türde, sebasöz ve apokrin bezleri, baskın ter bezleridir ve kıl köklerine yakın bir yerde bulunur. Bu bezlerin salgıları, kılları; yağlı, bazen köpüren bir karışım haline getirir (bir yarış atının koştuğunda ter içinde kalmasını düşünün). Bu tip bir terleme hayvanın serinlemesine yardımcı olur. Fakat ısının dağılarak yok olma yetisi sınırlıdır. University of Iowa’dan araştırmacıların yaklaşık 20 yıl önce ulaştığı bir sonuca göre, hayvanın derisi ıslandıkça ve bu kalın yağlı terle kaplandıkça serinleme etkisi azalıyor. Etkinliğin yok olması giderek artar çünkü derinin yüzeyinde değil, kürkün yüzeyinde buharlaşma meydana gelir, bu da ısı transferini engeller. Kürklü memeliler, uzun süreli egzersiz yapmaya zorlandıklarında, hipertermi (ısı bitkinliği) yaşar ve çökerler.
Kürkü olmamasının yanı sıra, insanlar, olağanüstü sayıda ekrin bezi sahibidir. Bu sayı, 2 milyon ile 5 milyon arasında değişkenlik gösterir ve günde 12 litreye kadar ince, sulu ter üretilebilmesini sağlar. Ekrin bezleri, kıl kökleri yakınında kümelenmez, bunun yerine deri yüzeyine görece yakın bir yerde bulunur ve küçük porlar (delikler) aracılığıyla ter boşaltımı yapar. Çıplak derinin ve sulu terin bu kombinasyonu, terin direkt olarak kürkte birikmesinden ziyade doğrudan çıplak deride birikmesine sebep olur ve insanların aşırı ısınmasını engeller. Bu soğutucu sistemimiz sayesinde, sıcak bir günde yapılacak bir maratonda, atlara üstünlük sağlayabiliriz.
İnsanlar, kürkü olmayan tek primat türü ve fazlasıyla ekrin bezi sahibi olmasından kaynaklı, en yakın akrabamız olan şempanzelerle ayrıldığımız yer olan insansı soyumuzdan beri çıplak ve terli derinin ortaya çıkmasına sebep olacak bir şey olmuş olmalı. Belki de bu dönüşüm, ilkim değişimiyle birlikte başladı. Hayvan ve bitki fosilleri kullanılıp antik ekolojik koşullar yeniden canlandırılarak, bilim insanları, yaklaşık 3 milyon yıl önce başlayan ve Dünya’nın, insan atalarımızın yaşadığı Doğu ve Orta Afrika’da kurutucu etki yapan bir küresel ısınma fazına girdiğini belirledi. Düzenli yağışlardaki bu azalmayla birlikte, açık otlak alanlar sunan ağaçlık çevreler, eski insansılar tarafından tercih edildi ve atalarımızın beslenme açısından yetindiği yiyecekler –meyveler, yapraklar, kökler ve tohumlar– nadir bulunmaya başladı ve tıpkı tatlı su kaynaklarında olduğu gibi mevsimsel açıdan da erişilebilirliği azaldı.
Kaynakların bu küçülmesine cevaben, atalarımız nispeten aceleci olan beslenme alışkanlıklarını terk etmek zorunda kaldılar. Yalnızca su bulmak ve yeterli kalori elde edebilmek için mütemadiyen aktif bir yaşam biçimine girdiler, dahası su ve yenilebilir bitkiler bulmak için uzak mesafelere gitmek durumunda kaldılar. Yaklaşık 2.6 milyon yıl önceye ait arkeolojik kayıtlarda, hayvan kemiklerinin ve taş aletlerin ortaya çıkmasıyla, insansıların bu zamanlarda beslenmelerine eti de katmaya başladıkları anlaşıldı. Hayvansal besinler, bitkisel besinlere kıyasla kalori bakımından bir hayli zengindi, fakat arazide daha nadir bulunuyordu. Dolayısıyla, etçil hayvanlar yeterli miktarda yiyecek temin edebilmek için otçullardan daha geniş ve daha uzak arazilerde dolaşması gerekiyordu. Av hayvanları aynı zamanda hareket eden hedeflerdi ve avın korunması da gerekiyordu, bu da avcılar açısından eti elde edebilmek için daha fazla enerji harcamak anlamına geliyordu.
İnsan avcıları ve leş yiyiciler açısından ise, doğal seçilim, zamanlarının bir kısmını ağaçlarda geçiren australopitek türü insansıları koşmaya ve uzun adımlarla yürümeye elverişli uzun bacaklılar haline dönüştürdü. (Bu modern form aynı zamanda, atalarımızı zaman zaman akşam yemeği olmaktan da kurtardı.) Ancak bu artan etkinlik düzeyinin bir bedeli vardı: Aşırı ısınma riskinin çok artması. 1980’li yıllardan başlayarak, İngiltere’deki Liverpool John Moores Üniversitesi’nden Peter Wheeler, sıcaklayan atalarımızın savanadan nasıl çıktıklarını canlandırdığı bir dizi makale yayımladı. Wheeler’ın çalışması, meslektaşlarımla beraber 1994’de yayınladığımız araştırmalarla birlikte şunu ortaya koyuyor: Kas aktivitesinin içsel olarak ısı kazandığı yürüyüş ve koşu sırasında geçirilen zaman arttıkça, hominidler hem ekrin terleme becerilerini arttırdı, hem de vücut kıllarını kaybetti. Böylece aşırı ısınmaktan kaçınabildiler.

Peki acaba bu dönüşüm ne zaman meydana geldi? İnsan fosil kayıtları cildi korumasa da araştırmacıların, atalarımızın çağdaş hareket şekillerine ne zaman başladıklarına dair kaba bir fikri var. Johns Hopkins Üniversitesi’nden Christopher Ruff ile Lieberman tarafından bağımsız olarak yapılan araştırmalar, yaklaşık 1.6 milyon yıl önce cinsimizin Homo ergaster adı verilen ilkel bir üyesinin, modern bir vücut oranına sahip olduğunu ve bunun da uzun yürüyüşe ve koşmaya izin vereceğini ortaya koymuştur. Üstelik bileğin, dizin ve kalçanın eklem yüzeylerinin ayrıntıları, bu hominidlerin aslında kendilerini bu şekilde etkilediğini açıkça ortaya koymaktadır. Dolayısıyla, fosil kayıtlarına göre, çıplak deriye ve ekrin temelli bir terleme sistemine geçiş, atalarımızın yeni yorucu yaşam biçimine eşlik eden daha büyük ısı yüklerini dengelemek için 1,6 milyon yıl önce ortaya çıkmaya başlamış olmalıdır.
Hominidlerin çıplak deriyi ne zaman geliştirdiğine ilişkin bir diğer ipucu, cilt renginin genetiği araştırmalarından gelmiştir. 2004’te yayınlanan ustaca bir araştırmada, Utah Üniversitesi’nden Alan R. Rogers ve meslektaşları cilt pigmentasyonu üreten genlerden biri olan insan MC1R gen dizilerini inceledi. Ekip, koyu pigmentasyonlu Afrika’lılarda hep rastlanan belli bir gen çeşidinin 1.2 milyon yıl önce ortaya çıkmış bulunduğunu gösterdi. Erken insan atalarının, şempanze gibi çok siyah kürkle kaplı pembe ciltleri olduğuna inanılıyor. Bu nedenle, kalıcı koyu cildin gelişimi, muhtemelen güneş koruyucu vücut kıllarının kaybedilmesini takiben oldu. Rogers’ın tahmini böylelikle kılsızlığın çıkışı için asgari bir tarih sağlıyor.

Çıplak deri, atalarımızın yaşadığı aşırı sıcak iklimlerde, insanların, normal vücut sıcaklığını korumak üzere evrimleştiği tek adaptasyon değildir. Aynı zamanda da, daha uzun uzuvlar geliştirmiş ve böylelikle aşırı ısıdan kurtulmayı kolaylaştıran yüzey-hacim oranını arttırmıştır. Bu eğilim, bugün de devam ediyor gibi görünüyor. Devam eden bu adaptasyona dair en iyi kanıt ise, Sudan’da yaşayan bir etnik grup olan Dinka gibi Doğu Afrika populasyonlarından geliyor. Yeryüzündeki en sıcak yerlerden birisinde yaşayan bu insanların son derece uzun uzuvlarının olması kesinlikle tesadüf değildir. Peki, modern insanlar neden bu denli geniş ölçülerde değişiklik gösteren uzuv oranlarını geliştiriyor? Atalarımız, tropikal Afrika’dan dünyanın daha serin bölgelerine göç ettikçe, seçilim baskıları da çeşitli vücut tiplerinin evrimleşmesini ortaya çıkaracak şekilde değişti.
Son yıllarda, araştırmacılar, insanların vücut kıllarını neden kaybettiklerine dair deliller bulabilmek amacıyla bitlere odaklandı. 2003 yılında University of Reading’den Mark Pagel ve John Radcliffe Hastanesi’nden Walter Bodmer; insanların, derilerindeki hastalık yayıcı bitlerden ve kıllara yerleşen diğer parazitlerden kurtulmak ve deri sağlığını korumak için kıllarını döktüklerini ileri sürdü. Diğer araştırmacılar ise, atalarımızın çıplak olmalarından ne kadar süre sonra “kıyafet” giymeye başladıklarına dair fikir sahibi olmak için kafa ve vücut bitleri üzerinde çalışmalar yürüttü. Her ne kadar, vücut bitleri kandan beslense de, kıyafetler üzerinde de yaşayabilir. Böylece, vücut bitlerinin kökeni, insansıların kıyafet anlayışlarının ortaya çıkmasına dair minimum bir tahmin sağlıyor. Organizmaların gen dizilimlerini karşılaştırarak, araştırmacılar, türlerin ne zaman ortaya çıktığına dair kabaca bir tahmin geliştirebilir. Bitlerde yürütülen böylesi analizler, kafa bitlerinin insanlarda başlangıçtan beri bulunduğunu ancak vücut bitlerinin daha sonra evrimleştiğini ortaya koyuyor. Bu türlerin ortaya çıkış zamanları, insanların “giyinmelerinden” yaklaşık bir milyon yıldan daha fazla bir süre önce çıplak olduklarına işaret ediyor.
‘İnsanımsılarda çıplaklık nasıl evrimleşti?’ sorusunun cevabı ‘Neden ve ne zaman vücudumunuz çıplaklaştı?’ sorusunun cevabından daha az biliniyor. Çıplaklığın evriminin genetik delillerinin belirlenmesi, derimizin görüntüsüne ve fonksiyonuna katkı yapan birçok gen olduğundan dolayı oldukça zor. Yine de, farklı organizmaların bütün genomlarındaki DNA kod harflerinin ya da nükleotidlerin büyük ölçekli karşılaştırmalarından, bir takım ip uçları elde edilebilir. İnsan ve şempanze genomlarının karşılaştırılması, şempanze DNA’sı ile bizim DNA’mız arasındaki en önemli farklılığın derinin özelliklerini kontrol eden proteinlerin kodlarında olduğunu açığa çıkartıyor. Bu genlerin bazılarının insan versiyonları, derimizi özellikle su geçirmez ve çizilmeye dayanıklı yapmaya yardımcı olan proteinleri kodluyor. Bunlar koruyucu kılların yokluğunda deride bulunması gereken kritik özellikler. Belirli gen değişkenlerinin gelmesinin çıplaklığın sonuçlarını hafifleterek çıplaklığa katkı yaptığı önermesi bu bulgular üzerinden yapılıyor.
Derimizin üstün engelleme kapasitesi, epidermisin stratum corneum (SC) olarak adlandırılan en dış katmanının yapısından ve karakterinden kaynaklanıyor. SC, binalardaki tuğla ve sıva yapısı gibi tanımlanabilir. Bu düzen içerisinde, korneositler olarak adlandırılan ve kretin proteini ile diğer maddeleri içeren düz ve ölü hücrelerin çoklu katmanlarını tuğlalara benzetebiliriz. Korneositlerin her birinin etrafını saran ultra-ince lipid katmanlarını da sıvaya.
SC gelişimini yönlendiren genlerin çoğunluğunun antik olduğu söylenebilir. Ayrıca bu genlerin dizileri, omurgalılarda oldukça iyi korunmuş durumda. İnsan SC’sini kuvvetlendiren genlerin belirgin işaretleri var, bundan dolayı bu genler hayatta kalmak için öneme sahip. Bu genler, yalnızca epidermis içerisinde meydana gelen ve keratin ile involucrinin özgün tiplerinin de dahil olduğu proteinlerin eşsiz kombinasyonlarının üretimini kodluyor. Birçok laboratuvarda, bu proteinlerin üretimini düzenlemekten sorumlu belirli mekanizmalar çözümlenmeye çalışılıyor.
Diğer araştırmacılar, insan derisi üzerindeki kılların seyrekliği ve inceliğinden sorumlu mekanizmaları belirlemek amacıyla vücut kıllarındaki keratinlerin evrimini araştırıyorlar. İlgili konu üzerinde yapılan araştırmalardan birisinde, Marburg’daki Philipps University’den Roland Moll ve çalışma arkadaşları, vücut kıllarındaki keratinlerin oldukça hassas olduklarını gösterdiler. Bu sebeple, insan vücudundaki kıllar diğer hayvanlarınkilere kıyasla daha kolay koparılabiliyor ya da kırılabiliyor. 2008 yılında yayımlanan bu çalışmanın önermesine göre, insan kıl keratinleri, evrim süresince diğer primatlarınki kadar hayatta kalmak için önemli değildi ve bu sebeple zayıfladılar.
Genetikçiler ayrıca, insan derisinin nasıl bu kadar bol ekrin bezi içerdiği sorusunu da cevaplamaya çalışıyorlar. Epidermal kök hücrelerin kaderini belirleyen genlerdeki değişikliklerden dolayı, bu birikmenin meydana geldiği neredeyse kesin. Gelişimin erken aşamalarında, belirli bölgelerdeki epidermal kök hücre grupları alt derinin altındaki hücreler ile etkileşime girerler ve bu hücreler içerisine genetik olarak iletilen kimyasal sinyaller kök hücrelerin kıl folüküllerine, ekrin bezlerine, apokrin bezlerine, yağ bezlerine ya da sade epidermise farklılaşmasını yönlendirir. Birçok araştırma grubu epidermal kök hücrelerin nasıl belirlendiğini ve korunduğunu araştırıyor. Bu çalışmalar embriyonik epidermal hücrelerin kaderini neyin yönlendirdiğini ve bu hücrelerin insanlarda ekrin ter bezlerine nasıl dönüştüğünü açığa çıkartacak.
Yine de çıplak apelere dönüşmüş olsak da, evrim vücudumuzun bazı parçalarını kıllarla kaplı bıraktı. Dolayısıyla insanların kürklerini neden kaybettiklerine dair herhangi bir açıklama, neden bazı bölgelerde hala biraz kaldığının da hesabını verebilmeli. Koltuk altlarındaki ve kasıklardaki kıllar muhtemelen hem feromon (diğer bireylerden davranışsal bir tepki almaya yarayan kimyasallar) salınımına yarıyor, hem de bu bölgelerin hareket sırasında nemli kalmasına yardımcı oluyor. Kafadaki saçlar ise, büyük olasılıkla başın üst kısmında meydana gelecek aşırı ısınmaya kalkan görevi görüyor. Bu düşünce kulağa mantıksız gelebilir; fakat başın üstünde yoğun kıllara sahip olmak, saçların sıcak yüzeyi ile terleyen kafa derisi arasında bir hava bariyer katmanı meydana getirir. Böylece sıcak ve güneşli bir günde, saçlar ısıyı absorbe ederken, hava bariyer katmanı serin kalmaya devam eder ve kafa derisinin terleyerek, terin bu hava katmanına doğru buharlaşmasına olanak sağlar. Sıkı ve kıvırcık saçlar bu bağlamda en uygun baş korumasını sağlar, çünkü saçların yüzeyi ile kafa derisi arasındaki boşluğun kalınlığını arttırarak, havanın bu boşluğa girmesine izin verir. İnsanlarda saçın evrimiyle alakalı daha keşfedilmemiş pek çok şey var; fakat sıkı kıvırcık saçlara sahip olmanın, modern insanların başlangıçtaki durumu olması ve diğer saç türlerinin insanlar tropik Afrika bölgesinin dışına doğru yayılırken evrimleşmiş olması mümkün.
Söz konusu vücut kıllarımız olduğunda ise, soru, neden bu kadar değişkenlik gösterdikleri haline dönüşüyor. Tüm vücudunda çok az kıl ve kıl bulunan üyelere sahip toplumlar olduğu kadar, ayrıca vücudu tüy ve kıllarla kaplı bireylere sahip toplumlar da var. Vücudu çok az kılla kaplı olanlar tropik bölgelerde yaşamaya eğilimliyken, vücudunda çok fazla kıl bulunanlar tropik bölgeler dışında yaşamaya eğilimli. Yine de tropik olmayan bölgelerde yaşayan insanlardaki bu vücut kılları, kayda değer bir sıcaklık sağlamıyor.
Kıllılıktaki bu değişkenlik, kesinlikle testosteron miktarından kaynaklanıyor, çünkü bütün toplumlarda erkekler, kadınlara kıyasla daha fazla vücut kılına sahip. Bu dengesizliği açıklamayı konu edinen bazı teoriler ise, bunu cinsel seçilime bağlıyorlar. Örneğin bu teorilerden birisinde, kadınların daha gür sakallı ve daha kalın vücut kıllarına sahip erkekleri tercih ettikleri, çünkü bu özelliklerin güç ve cinsel iktidar ile beraber meydana geldiği öne sürülüyor. Bir diğeri ise; erkeklerin, kadınlarda daha çocuksu özelliklere dair bir tercih geliştirdiğini iddia ediyor. Bunlar ilginç hipotezler, fakat henüz kimse bunları modern insan nüfusu üzerinde sınamadı; bu nedenle, örneğin kıllı erkeklerin, daha köse erkeklere kıyasla, gerçekten daha güçlü ve doğurgan olup olmadıklarını bilmiyoruz. Deneysel kanıtların yokluğunda, insanlardaki vücut kıllarının neden şu anki haliyle değişkenlik gösterdiği, hala birilerinin tahmininden ibaret.
Evrimsel süreçte kılların kaybolması elbette her şeyin son bulması anlamına gelmiyor ancak etkileri hiç de azımsanacak cinsten değil. Şöyle ki; kılsızlaşmayı takip eden süreçteki insan evrimi fazlarında bu sürecin çok güçlü sonuçları oldu. Vücut kıllarının büyük bir bölümünü kaybetmek, fazla vücut ısısının ekrin bezlerinden terleme ile yayılmasını ve böylelikle ısıya en duyarlı olan organımızın yani ‘beynimizin’ bugün sahip olduğumuz efektif ve büyük haline gelmesini mümkün hale getirdi.
Australopitekler, modern şempanzeninkine yakın büyüklükte 400 cm3`lük bir beyne sahipken, Homo ergaster bunun neredeyse iki katı büyüklüğünde bir beyne sahipti. Homo ergasteri takiben bir milyon yıllık evrimsel süreçte insan beyni ekstra bir 400 cm3 hacim kazanarak bugünkü modern büyüklüğüne ulaştı. Elbette modern beynin oluşumunu sağlayan tüm evrimsel süreçler bundan ibaret değil. Örneğin, gri madde dokusunun ve beyni kaplaması kıvrımların oluşumunu uyaran tüm genetik ve çevresel faktörler de bugünkü beynin işlevsel konumun açıklanmasında büyük bir önem arz ediyor. Ancak, vücut kıllarının azalmasının da gelişkin bir beyne sahip olmanın önünü açtığı ve evrimsel olarak önemli büyük bir adımın atılmasını kolaylaştırdığı şu an için mevcut şüphelerin üzerinde kabul edilmiştir denilebilir.
Bununla birlikte ‘kılsızlığımız’ın sosyal bir takım yansımaları da süregelmiştir. Duygu-durumsal konumumuza göre tüy kökleri veya kıl foliküllerinin diplerinde bulunan küçük kasların kasılıp gevşemesi istemsizce yaptığımız hali ile tüylerimizin diken diken olmasını, ürpermesini sağlamaktadır. Bu noktada söylemek gerekir ki, tüylerimizle duygularımızı göstermekte çok da iyi sayılmayız. Çünkü kaybettiğimiz kıllar ve kıl yoğunluğu; kediler ve köpekler gibi tüylü canlıların aksine tüylerimiz ince ve zayıf olduğundan duygularımızı bu yolla göstermemizin zorlaşmasına sebep olmuştur. Kılsızlık, yine zebralar gibi kıllarımız üzerinden kendimizi tanıtacak karakterlere sahip olamamamıza kısacası kıllarımız üzerinden iletişim kuramamamıza yol açmıştır. Bu durum, bir grup bilim insanının yüz ifadelerimizi geliştirmemizin çokça jest ve mimik kullanmamızın ve hatta kompleks dil yapıları geliştirmemizin sebeplerinden birinin ‘kürksüz’ canlılar olmamız olarak öne sürmelerinin alt metnini oluşturmuştur.
Bugün kullandığımız kozmetik ürünlerinin, vücut boyalarının hatta belki solaryumun, daha da ileri gidecek olursak kıyafetlerin ve modanın bile benzer bir karakter eksikliğini kapatma ihtiyacı ile ortaya çıkmış olması kuvvetle muhtemeldir. Bütüne bakıldığında, kürkümüzü kaybetmek bizi yalnızca daha serin, gelişmiş beyinli değil; ‘insan’ yapan şeydir diyebiliriz.
Orjinal yazı: Bilimfili; https://bilimfili.com/ciplak-gercek-insanlarin-kurku-yoktur/
Kaynak&İleri Okuma:


