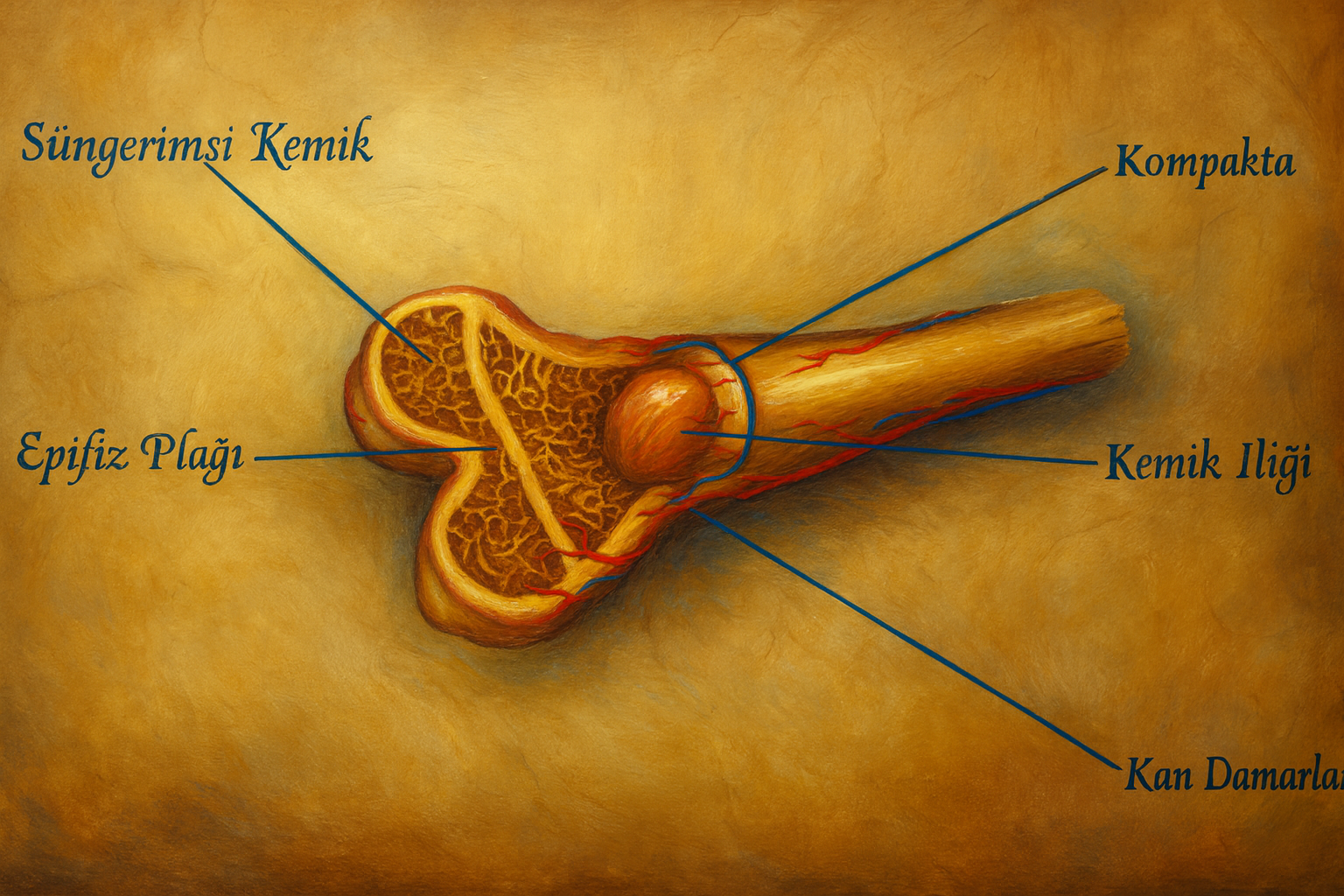
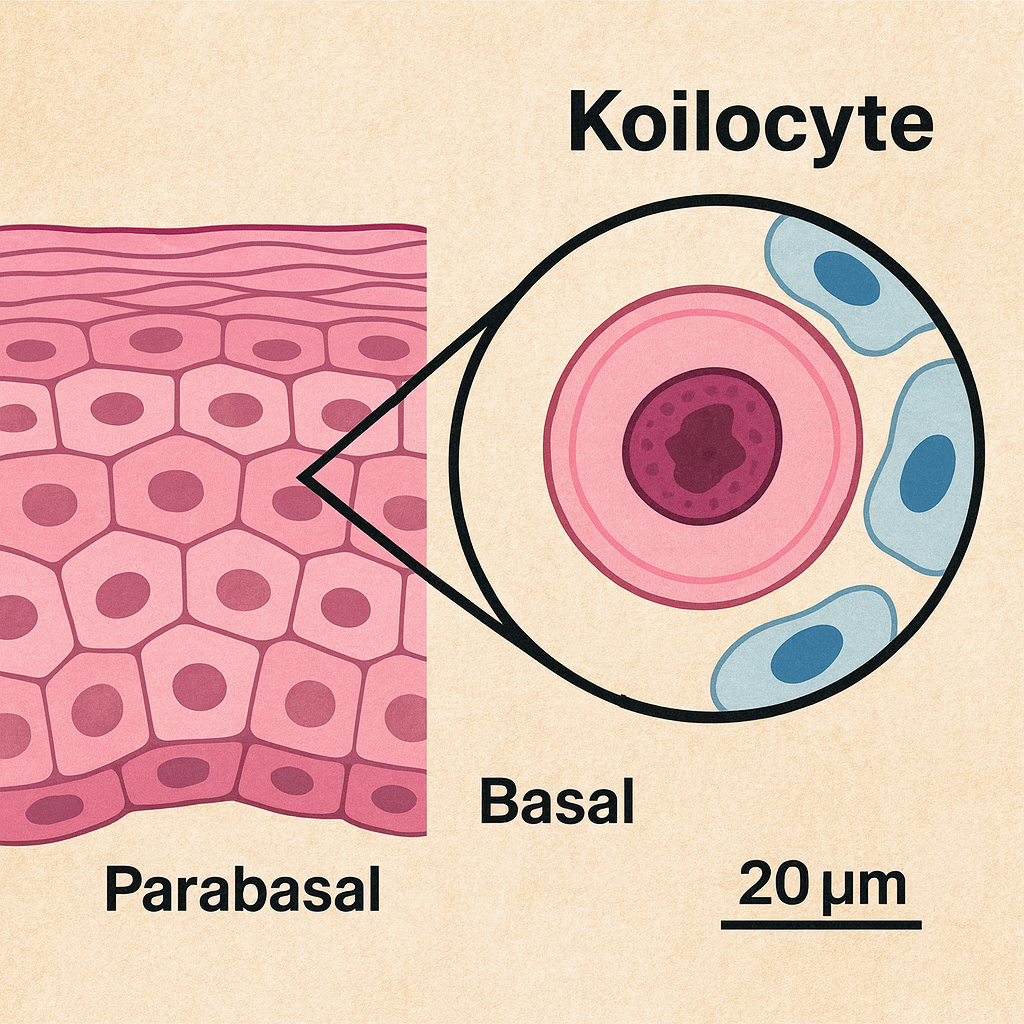
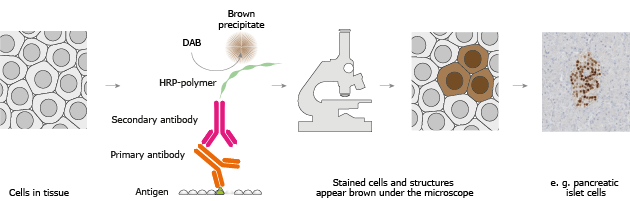
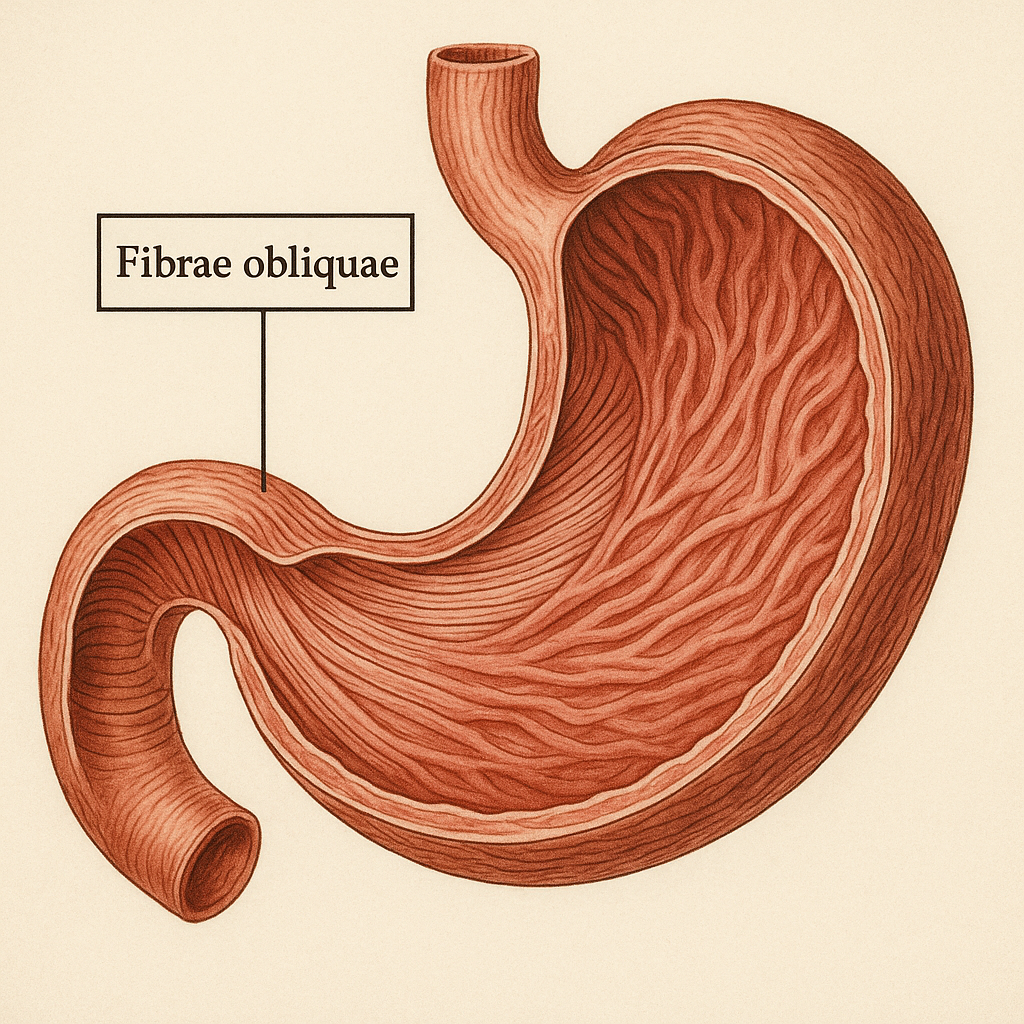
Azurofilik
“Azurofilik” terimi, mavinin bir tonu olan “azure” sözcüğünden ve “güçlü bir yakınlığa sahip olmak” anlamına gelen Yunanca “philic” teriminden türetilmiştir. Tarihsel olarak azurofilik, mikroskobik boyama tekniklerindeki gelişmelerle, özellikle de gök mavisi boyaların histoloji ve hücresel biyolojide yaygın olarak kullanılmaya başlanmasıyla ön plana çıkan bir terimdi. Azurofilik, belirli hücre yapılarının gök mavisi boyalara bağlanma eğilimini ifade … Devamını oku