İçindekiler
Etimoloji ve kavramsal çerçeve
“Limbik” sözcüğü, Latince limbus (“kenar, sınır”) kökünden, Fransızca limbique aracılığıyla türemiştir ve serebral hemisferlerin iç yüzünde, corpus callosum’un çevresini ve medial temporal lobu kıyılayan yapılar bütününe işaret eder. Filogenetik açıdan neokorteksten daha eski (allo-/mezo-korteks) alanları yoğun biçimde içerir; ancak güncel nörobilim, limbik sistemin katı sınırlarla ayrılmış tekil bir “modül” olmaktan ziyade, kısmen örtüşen ağlar kümesi olduğunu vurgular. Dürtüsel davranışların ayarlanması, duygulanım, güdülenme, öğrenme ve bellek süreçleri ile otonom-endokrin düzenleme (yeme, sindirim, üreme davranışları ve stres yanıtı gibi) üzerinde merkezi bir etkisi vardır. Bununla birlikte bu işlevlerin beyin genelinde dağılmış ağlarca üretildiği, limbik yapıların ise bu ağlar içinde düğüm (“hub”) görevi gördüğü özellikle not edilmelidir.
“Limbik sistem” terimi tarihsel ve öğretici değeri yüksek olmakla birlikte, nörobiyolojik doğruluk açısından “limbik ağlar” ifadesi daha uygundur. Sınırlar bağlamsal olup çalışmanın yöntemine (lezyon, doku boyama, DTI, fMRI, elektrofizyoloji) göre değişebilir.
Gelişimsel ve filogenetik özellikler
Limbik yapılarda üç tabakalı arşikorteks (hipokampal oluşum) ve beş tabakalı paleokorteks (ör. entorhinal ve perirhinal alanların kısımları) belirgindir; bu korteksler altı tabakalı neokortekse kıyasla filogenetik olarak daha eskidir. Bu histoarkitektur, limbik ağların yüksek plastisite, güçlü iç tekrarlayıcı bağlantılar ve yaygın nöromodülatör (kolinerjik, noradrenerjik, dopaminerjik) girdilerle karakterize olmasına zemin hazırlar.
Anatomik kapsam ve sınırlar
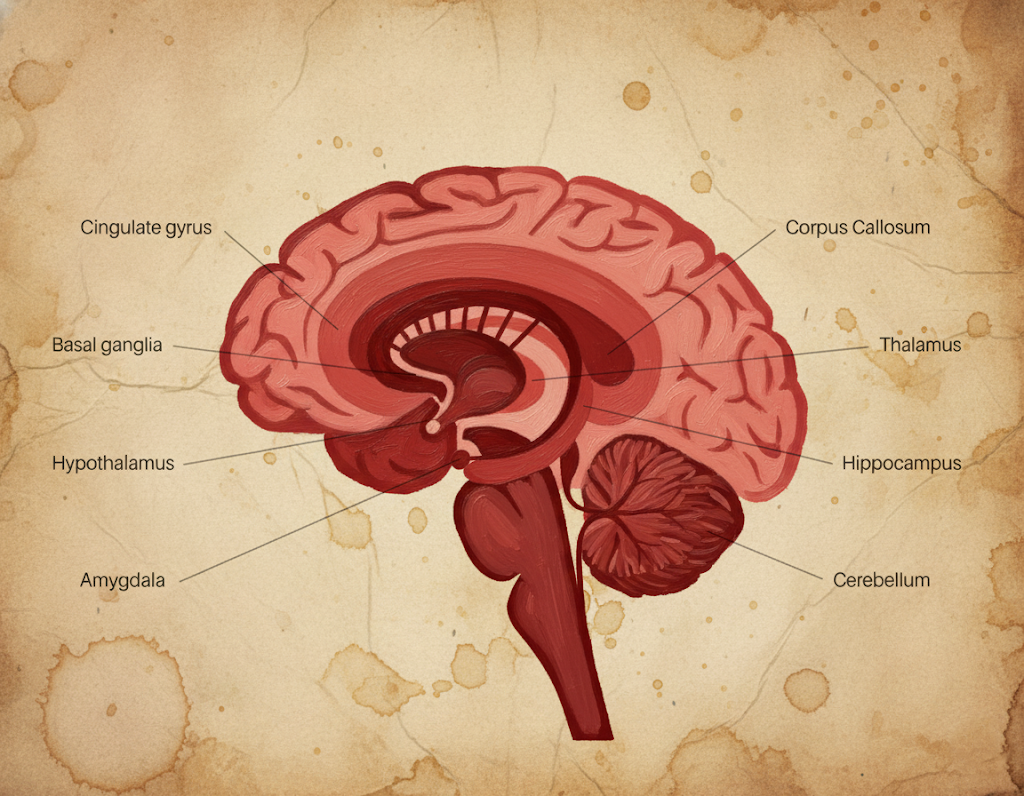
Klasik “limbik lob” tanımı medial frontal ve parietal yüzeyde gyrus cinguli ile medial temporal lobda gyrus parahippocampalis’i, bunların altındaki hipokampal oluşumu ve bunlarla sıkı bağlantılı derin çekirdek ve traktları içerir. Aşağıda, kullanıcı tarafından belirtilen başlıca yapılar sistematik biçimde özetlenmiştir.
Corpora mamillaria (Mamiller cisimler)
- Konum ve bağlantılar: Hipotalamusun arka-alt bölümünde, üçüncü ventrikülün tabanına komşu çift çekirdek kompleksidir. Başlıca afferentlerini forniks aracılığıyla hipokampustan alır; efferentleri mamillotalamik trakt üzerinden anterior talamik çekirdeklere ve oradan singulat girusa iletilir.
- İşlev: Epizodik bellek devrelerinde röle konumundadır.
- Klinik: Mamiller cisim/ mamillotalamik trakt lezyonları ağır anterograd amneziye yol açabilir; kronik tiamin eksikliği (Wernicke–Korsakoff spektrumu) seçici duyarlılık gösterir.
Fornix cerebri (Forniks)
- Makroanatomi: Hipokampus alveusundan çıkan liflerin fimbria üzerinden birleşmesiyle oluşan, krus–gövde–kolonlara ayrılan C-şekilli ana çıkış traktıdır.
- Bağlantı mantığı: Postkomissüral lifler mamiller cisimlere; prekomissüral lifler septal alanlara ve ventral striatal hedeflere uzanır.
- Klinik: Forniks kesileri, anevrizma/kraniyotomi sonrası hasarlar ve üçüncü ventrikül cerrahileri epizodik bellek kusuruna yol açabilir.
Gyrus cinguli (Singulat girus)
- Alt bölümler: Rostral/ventral anterior singulat (duygulanım–otonom entegrasyon), dorsal anterior singulat (bilişsel kontrol–hata izleme), posterior singulat ve precuneus (otobiyografik bellek–içsel yönelimli dikkat).
- Bağlantılar: Cingulum demeti aracılığıyla prefrontal, parietal ve parahipokampal alanlarla geniş iki yönlü bağlantılar.
- İşlev: Duygulanımın bilişsel kontrolle birleştiği “arka plan düzenleyicisi”; ağrı deneyiminin duyuşsal bileşeni; hedefe yönelik davranışların sürdürülmesi.
Gyrus parahippocampalis (Parahipokampal girus)
- Bileşenler: Entorhinal korteks (BA 28), perirhinal (BA 35/36) ve parahipokampal korteks; hipokampal oluşumun majör neokortikal kapısıdır.
- İşlev: Bağlamsal/çevresel sahne işleme ve epizodik bellek indeksleme; entorhinal ızgara hücreleri uzamsal haritalamaya katkı verir.
Hippocampus (Hipokampus ve hipokampal oluşum)
- Mikrodevre: Trispatik çevrim: entorhinal → dentat girus → CA3 (kollateral tekrarlı ağ) → CA1 → subikulum/entorhinal geri besleme.
- Eksensel organizasyon: Anterior (ventral) kısım duygulanım ve stresle, posterior (dorsal) kısım ayrıntılı uzamsal/epizodik temsil ile daha çok ilişkilendirilir.
- Fizyoloji: Uzun süreli güçlenme (LTP), “yer hücreleri” ve hızlı örnekleme temelli epizodik bağ kurma.
- Klinik: İki taraflı mediyal temporal hasar belirgin anterograd amnezi doğurur; Alzheimer hastalığında entorhinal–hipokampal ağ erken etkilenir; mezial temporal epilepside skleroz ve devre yeniden örgütlenmesi tipiktir.
Corpus amygdaloideum (Amigdala kompleksi)
- Nükleer mimari: Bazolateral kompleks (lateral, bazal, aksesuar-bazal), santral çekirdek, medial ve kortikal gruplar.
- Girdiler/Çıkışlar: Duyusal birlikteliği yüksek çok modlu afferentler (özellikle temporal korteks); efferentler stria terminalis ve ventral amigdalofugal yol üzerinden hipotalamus, BST, beyin sapı ve orbitofrontal/medial prefrontal alanlara.
- İşlev: Tehdit algısı ve koşullanmış korku, değer-atama (salience), sosyal-olgusal ipuçlarının hızla bütünleştirilmesi.
- Klinik: Lezyonlarda korku tanıma/koşullanması zayıflar; aşırı reaktivite travma ile ilişkili bozukluklarda ve anksiyete spektrumunda görülür.
Indusium griseum (İndusium grisium; suprakallozal girus)
- Tanım: Corpus callosum’un üst yüzeyini örten ince gri cevher tabakası; filogenetik olarak hipokampal oluşumun kallozal yüzeye uzanan kalıntısıdır.
- Bağlantılar: Longitudinal strialar (Lancisi çizgileri) boyunca seyreden ince liflerle komşu limbik alanlara sınırlı bağlantılar.
- Not: Erişkin insan beyninde işlevsel katkısı görece mütevazı ve çoğu kaynakta vestigial kabul edilir.
Subiculum (Subikulum)
- Konum: Hipokampus CA1 ile entorhinal korteks arasında geçiş (transisyonel) korteks; hipokampusun ana çıkış kapısıdır.
- Bağlantılar: Fornix, anterior talamus, retrosplenial/singulat ve ventral striatal hedeflere yaygın projeksiyonlar.
- İşlev: Epizodik bağlamsallaştırma, yer–yön dönüşümü ve hipokampal çıktının ağlar arası dağıtımı.
Nucleus interpeduncularis (İterpedinküler çekirdek)
- Konum: Orta beynin orta hatta, pedinküller arası fossada yer alan tegmental çekirdek kompleksi.
- Bağlantılar: Üstten habenuladan fasciculus retroflexus ile yoğun girdi alır; limbik telensefalik kaynakların (özellikle septo-hipokampal ve amigdalar projeksiyonların) habenular röle üzerinden beyin sapına aktarımında önemli bir düğümdür.
- İşlev/Klinik: Aversif pekiştirme ve otonom-refleks düzenlemelerde rol; nikotin bağımlılığı/çekilme fizyolojisinde ve duygu-durum düzenlemelerinde habenulo-tegmental devrelerin parçası olarak incelenmiştir.
Bağlantısal organizasyon: ana demet ve devreler
- Papez devresi: Hipokampus → (fimbria/fornix) → mamiller cisimler → (mamillotalamik trakt) → anterior talamus → singulat girus → (cingulum) → entorhinal/parahipokampal → hipokampus. Epizodik bellek ve duygu-bellek etkileşimi için iskelet bir halka.
- Amigdalar ağları: Stria terminalis ve ventral amigdalofugal yol ile hipotalamus, BST ve beyin sapına; bazolateral kompleks üzerinden orbitofrontal/medial prefrontal kortekse değer-atama ve karar devreleri.
- Septo-hipokampal devre: Medial septum–hipokampus arası iki yönlü bağlantılar hipokampal teta ritmini ve keşif/öğrenme dinamiklerini ayarlar.
- Cingulum demeti: Singulat–retrosplenial–parahipokampal eksende uzunlamasına lifler; epizodik yönlendirme ve içsel gezinme (mental navigation) için taşıyıcı bant.
- Habenulo–interpedinküler sistem: Limbik ön beyin kaynaklarını aversif/ödüllendirici sonuç sinyallerine dönüştürerek beyin sapı nöromodülatör çekirdeklerini eğler.
Temel işlevsel katkılar
Duygulanım ve güdülenme
Amigdala ve ventromedial/ orbitofrontal ön korteks arasındaki döngüler, uyaranların biyolojik önemini (salience) ve duygusal tonunu hızla belirler. Anterior singulat, bu değerlendirmeleri çatışma izleme, hata sinyali üretimi ve otonom yanıtlarla bütünleştirir. Hipotalamus aracılığıyla otonom–endokrin çıktılar somatik duruş (visseral işaretler) olarak beden durumuna yansır.
Öğrenme ve bellek
Hipokampus, epizodik/bağlamsal temsilleri birkaç saniyeden yıllara uzanan sürelerde indeksler; parahipokampal- perirhinal ağlar sahne ve nesne/kavramsal birliktelikleri sağlar. LTP/LTD gibi sinaptik plastisite mekanizmaları ve “yer/ızgara/baş-yön” hücreleri uzamsal bellek ve epizodik yeniden çağırmayı destekler. Mamillotalamik eksen ve retrosplenial–anterior singulat alanları, anıların yön-bütünlüğünü koruyan talamo-kortikal rölelerdir.
Otonom ve homeostatik entegrasyon
Limbik yapılar, hipotalamus üzerinden enerji dengesi, besin alımı ve üreme davranışlarını bağlamsal hedeflerle ilişkilendirir. Amigdala ve hipokampus, öğrenilmiş ipuçlarıyla yeme davranışını (haz–ceza, anı–koku–mekân eşleşmeleri) şekillendirir; stres ekseni (HPA) aktivasyonu hipokampusta glukokortikoid-duyarlı plastisite değişimlerine yol açabilir.
Klinik-korelasyonel notlar
- Amnezik sendromlar: Hipokampus, forniks, mamillotalamik trakt ve anterior talamus boyunca herhangi bir halka kırılması belirgin epizodik bellek bozukluğu doğurabilir.
- Mezial temporal epilepsi: Hipokampal skleroz, amigdalo-hipokampal devrelerde yeniden örgütlenme; cerrahi rezeksiyonlarda bellek–dil dengelenmesi kritik önemdedir.
- Duygulanım bozuklukları ve travma: Amigdala hiperreaktivitesi, medial prefrontal–singulat hiporegulasyonu ve hipokampal hacim/bağlantısallık değişimleri; korku söndürme devrelerinde (infralimbik homologları) plastisite.
- Klüver–Bucy spektrumu: İki taraflı anterior temporal (amigdala ağırlıklı) lezyonlarda korku azalması, hiperseksüalite ve çevresel bağımlılık davranışları.
- Ağrı ve obsesif-kompulsif bozukluk cerrahisi: Singulotomi ve kapsulotomi gibi müdahaleler, limbik–fronto-striatal devrelerde hedefli modülasyon örnekleridir.
- Nörodejenerasyon: Alzheimer patolojisinin erken odakları entorhinal–hipokampal ağdır; retrosplenial ve posterior singulat hipo-metabolizması hastalığın işlevsel imzaları arasındadır.
Keşif
Broca’nın Paris’teki amfisinde 19. yüzyılın son çeyreğinde kullandığı büyük duvar çizimlerinde, corpus callosum’u sahilden geçen bir kıyı şeridi gibi kuşatan bir kıvrım şeridi belirir; Broca buna “grand lobe limbique” der ve Latince limbusun –kenar, sınır– çağrışımını tıp diline yerleştirir. Bu “kenar lobu”, gyrus cinguli’yi ve gyrus parahippocampalis’i bir halka gibi bağlar; halkanın iç yüzünde hipokampal oluşumun kemerli silueti seçilir. Öğrencilerinin notlarında, bu halkanın insan ve hayvan beyinlerinde kıyaslanabilirliğine yapılan vurgunun altı çizilidir: Filogenezde eski olanın morfolojisi, fonksiyonun ipucudur. Fornix’in mamiller cisimlere uzanan C-şekilli rotası, Broca’nın şemasında henüz bir “öykü” değildir; fakat çizgiler arasına saklanmış bir hareket hissi vardır: Hipokampustan çıkan lifler, sanki belleğin kendisi gibi, iz bırakarak ilerler.
20. yüzyılın ilk yarısında, Papez’in 1937 tarihli önerisi bu harekete bir ritim verir. Hipokampus → fornix → corpora mamillaria → mamillotalamik trakt → anterior talamus → gyrus cinguli → cingulum → parahipokampal girus → hipokampus döngüsü, duygulanımın “devresi” olarak bir araya getirilir. Papez’in metninde, anatomik bir haritanın detayı ile fizyolojik bir sezginin cüreti yan yanadır: Duygu, korteksin kıyısında doğar, talamusta yankılanır, singulatın kemerinde bir nitelik kazanır. Bu öneri, laboratuvardan çok anatomi salonunun kokusunu taşır; ama kapıları aralar. Anterior talamusun, epizodik bütünlüğün bekçisi gibi davranabileceği; gyrus cinguli’nin, otonom düzenleme ile bilişsel değer biçme arasında bir kavşak olabileceği fikri, klinisyenlerin ve deneycilerin zihninde yuva bulur.
Aynı yıllarda primat nörolojisinin sahnesinde, Klüver ve Bucy’nin iki taraflı anterior temporal rezeksiyonlarıyla davranış repertuarı sarsılan maymunlar belirir. Korku tepkilerinin sönmesi, hiperseksüalite, çevresel bağımlılık… En dramatik değişimler amigdala kompleksinin alanında yoğunlaşır. Corpus amygdaloideum, yalnızca bir anatomik çıkıntı değil, anlam yükleyen, tehdidi ve ödülü hızla derecelendiren bir düğüm olarak görünür olur. Stria terminalis ve ventral amigdalofugal yol boyunca hipotalamusa ve beyin sapına inen liflerin çizdiği hat, “visseral” olan ile “kognitif” olan arasında gerilimli bir iletken gibidir.
Savaş sonrası nöroanatomi, yalnızca boyaların değil metodolojinin de değiştiği bir dönemi başlatır. Nauta’nın lif izleme teknikleri, Heimer’in ventral striatal bağlantı şemaları, septo-hipokampal devrenin teta ritmiyle atıp tutan iki yönlü doğasını görünür kılar. Fornix artık yalnız bir köprü değil; postkomissüral kollarıyla corpora mamillaria’ya, prekomissüral lifleriyle septal ve ventral striatal hedeflere ayrışan akışlar dizgesidir. Mamillotalamik traktın ön talamusa ve oradan singulat girusa uzanan hattı, Papez’in sezgisel devresine anatomik bir omurga kazandırır. Bu arada, beyin sapının gölgeli orta hattında, nucleus interpeduncularis’in habenula ile kurduğu aks –fasciculus retroflexus boyunca– aversif pekiştirmenin ve nikotinerjik düzenlemenin ince ayarlı bir istasyonu olarak seçilmeye başlar; limbik telensefalonun telkini, tegmental çekirdeklerin ritmine karışır.
Klinik sahnede 1957’de H.M.’nin hikâyesi, limbik anlatının eksenini dramatik biçimde değiştirir. Mediyal temporal lob rezeksiyonunun ardından ortaya çıkan derin anterograd amnezi, hipokampusun ve komşu parahipokampal–entorhinal ağların epizodik bellekteki zorunlu payını tartışmasız kılar. Subikulumun –hipokampus CA1 ile entorhinal korteks arasında– bir “geçiş korteksi” olarak üstlendiği dağıtım rolü, klinik olguların ve deneysel hayvan çalışmalarının satır aralarında giderek daha belirginleşir. Aynı dönemde, tiamin eksikliğinin örselediği mamiller cisimler ve mamillotalamik trakt, dize gelen anlatıyı tamamlar: Bellek yalnızca bir yer değil, bir hatlar sistemidir; herhangi bir halkası koptuğunda öykü yarım kalır.
MacLean’in 1949’da “visseral beyin”, sonraki yıllarda “üçlü beyin” başlıklarıyla geniş kitlelere ulaşan çerçevesi, kavrama güçlü bir popüler ad verir: “limbik sistem.” Üç katmanlı evrimsel mimari fikri, ders amfilerinde kolay kavranan bir resim sunsa da, araştırma derinleştikçe bu resmin fazla düzgün olduğu anlaşılır. Limbik olanın çekirdeği –gyrus cinguli’nin kemeri, gyrus parahippocampalis’in sahnelemesi, hipokampal oluşumun arşikortikal laminaları, amigdalanın nükleer mozaği– birlikte çalışır; fakat her bağlamda aynı orkestrasyonla değil. İnsanın karmaşık toplumsal ve bilişsel ihtiyaçları, bu çekirdeği prefrontal alanların ve bazal gangliyon döngülerinin geniş bir karnavalına bağlar.
1970’lerle birlikte limbik fizioloji, mikrodüzeyde bir anlatıya kavuşur. O’Keefe’in hipokampusta “yer hücreleri”ni tanımlaması, Bliss ve Lømo’nun uzun süreli güçlenmeyi göstermesi, epizodik haritalamanın sinaptik izlerini gözle görünür kılar. Dentat girustan CA3’e, oradan CA1’e uzanan trispatik çevrim, tekrarlı kollaterallerin ve rekürrent dinamiklerin diliyle konuşur. Entorhinal korteksin ızgara hücreleri, yıllar sonra uzamsal metrik için bir zemin çizecektir; şimdilik, parahipokampal–perirhinal sahnelerin ve nesne bağdaştırmalarının kapısı olarak rolünü sürdürür. İndusium griseum bu büyük anlatıda sessiz bir kenar notudur: Corpus callosum’un sırtını örten ince gri örtü, antik gelişimsel katmanların yetişkin beyindeki mütevazı yankısıdır; longitudinal strialar boyunca geçen zarif lif demetleri, tarihsel hafızanın anatomiye düşürdüğü bir dipnottur.
1990’lardan itibaren amigdalanın deneysel psikolojideki yıldızı parlar. Koşullu korku, söndürme, genelleme; bazolateral kompleksin duyusal birlikteliği yüksek girdileri değerlendirip santral çekirdeğin hipotalamik–beyin sapı çıkışlarına ritim verdiği bir siyaset alanı gibidir. Anterior singulatın rostral bölümünde duygulanım–otonom entegrasyon, dorsal bölümünde hata izleme ve çaba tahsisi; posterior singulat–retrosplenial şeritte otobiyografik sahnelerin sessiz kurgusu… Cingulum demeti, bütün bu sahneler arasında bir yürüyüş yolu gibi uzanır. Bu yıllarda insan beyni kayıtları, derin elektrotların tek nöron düzeyindeki fısıltılarını alır; hipokampusun keskin dalga–ripple örüntüleri, uykuda ve uyanıklıkta anı tekrarlarının sessiz montajını yapar.
Yeni yüzyıla yaklaşırken, “ağlar çağı” başlar. Dinlenim durum ağı çalışmalarında posterior singulat–medial prefrontal–parahipokampal eksen, içe dönük düşünmenin ve epizodik dolaşmanın ana yollarını çizer; limbik–paralimbik kuşağın, varsayılan kiplik ağlarıyla iç içe geçmesi, “sistem” yerine “ağlar” dilini zorunlu kılar. Bağlantısal anatominin difüzyon tensör görüntüleme ile makro ölçekte, 7T fonksiyonel görüntüleme ile mezo ölçekte, laminar çözümlü kayıtlarla mikro ölçekte eşlenmesi; cingulumun uzunlamasına liflerinin, uncinat fasikülün orbitofronto–temporal diyaloğunun, fornixin komissür öncesi/sonrası ayrışmasının ayrıntılarını katman katman ortaya koyar. Ventral striatum–talamo–kortikal döngüler, “limbik” niyeti ödül beklentisi ve eylem seçimiyle birleştiren kapılar gibi çalışır.
Aynı dönemde klinik ve temel bilim birbirini yeniden besler. Subkallozal singulat hedefli derin beyin uyarımı, tedaviye dirençli depresyonda duygulanım ağlarına müdahalenin mümkün olduğunu gösterir; anterior singulotomi ve kapsulotomi, ağrı ve obsesif-kompulsif belirtilerde fronto–striatal–limbik hatların cerrahi yeniden ayarını örnekler. Mediyal temporal epilepsi cerrahisi, hipokampal sklerozun devre düzeyinde yeniden örgütlenmesini ve bellek–dil ekonomisinin kırılgan dengesini klinik anatomiye taşır. Mamillotalamik trakt hasarlarının “yönlü” bellek kusurlarına, anterior talamus lezyonlarının epizodik kurgudaki boşluklara nasıl denk düştüğüne dair vaka serileri, Papez’in şemasına çağdaş bir klinik gölge düşürür.
2010’lar ve 2020’lerde limbik hikâye, iki yeni motif kazanır. Birincisi, engram tartışmasıdır: Belirli nöron topluluklarının edinim ve geri çağırmada seçici olarak etkinleştirilip bastırılabilmesi, hipokampal–amigdalar düğümlerin belleğin hücresel anlatısına yerleşmesini sağlar. İkincisi, gradyanlar ve çokölçekli mimaridir: Duyusal alanlardan transmodal–içe dönük alanlara uzanan bağlantısal gradyanın en “içteki” ucu, limbik–paralimbik kuşağın üzerine düşer. Bu uçta, subikulum–entorhinal geçidin çokkatmanlı zaman–mekân kodları, anterior singulatın çaba–değer hesapları ve amigdalanın belirginlik (salience) atamaları, tek bir sistemden ziyade bir “ağlar orkestrasyonu” olarak çalışır. Nucleus interpeduncularis–habenula aksının aversif sonuç tahmininde üstlendiği ince ayar, dopaminerjik ve kolinerjik modülasyonun altyapısına duygu rengini katar; enerji dengesi ve stres biyolojisiyle çakışan bu hat, yeme davranışı ve kaçınma öğrenmesinin gölgeli yan sokaklarını aydınlatır.
Bugün, Broca’nın kıyı şeridi metaforu hâlâ geçerliliğini korur; ancak kıyıya vuran her dalga aynı denizden gelmez. Gyrus cinguli’nin kemerinden gyrus parahippocampalis’in sahnesine, hipokampus ve subikulumun geçitlerinden corpora mamillaria ve anterior talamusa, oradan da amigdalanın nükleer mozağine ve nucleus interpeduncularis’in orta hat istasyonuna uzanan yollar, bağlama göre farklı tempolarla akışa girer. Fornix bazen belleğin ağır adımlarıdır, bazen duygulanımın hızlı çağrışımları; indusium griseum sessizce, bir zamanlar daha geniş olanın artık ince bir iz olduğunu hatırlatır. “Limbik sistem” adı, bir müfrezenin değil, bir senfoninin başlığıdır; partisyonunda her yapı kendi temasını taşır ve çağın yöntemleri –intrakraniyal kayıtlar, yüksek alan görüntüleme, optogenetik ve hesaplamalı ağ bilimi– bu temaların hem birlikteliğini hem ayrışmasını duyulur kılar. Böylece keşif tarihi, sabit bir krokinin ilerletilmesi değil, yeni ışıkların altında aynı haritanın tekrar tekrar okunmasıdır: Her okumada, kenarın –limbusun– çizgisi biraz daha keskinleşir, fakat hiçbir zaman tek bir çizgiye indirgenmez.
İleri Okuma
- Broca, P. (1878). Anatomie comparée des circonvolutions cérébrales: Le grand lobe limbique et la scissure limbique dans la série des mammifères. Revue d’Anthropologie, 1, 385–498.
- Papez, J. W. (1937). A proposed mechanism of emotion. Archives of Neurology and Psychiatry, 38(4), 725–743.
- Kluver, H., & Bucy, P. C. (1939). Prefrontal lobotomy in chimpanzees. Journal of Neurophysiology, 2(5), 353–366.
- MacLean, P. D. (1949). Psychosomatic disease and the visceral brain; recent developments bearing on the Papez theory of emotion. Psychosomatic Medicine, 11(6), 338–353.
- MacLean, P. D. (1952). Some psychiatric implications of physiological studies on frontotemporal portion of limbic system (visceral brain). Electroencephalography and Clinical Neurophysiology, 4(4), 407–418.
- Scoville, W. B., & Milner, B. (1957). Loss of recent memory after bilateral hippocampal lesions. Journal of Neurology, Neurosurgery & Psychiatry, 20(1), 11–21.
- Nauta, W. J. H. (1958). Hippocampal projections and related neural pathways to the midbrain in the cat. Brain, 81(2), 319–340.
- O’Keefe, J., & Dostrovsky, J. (1971). The hippocampus as a spatial map: Preliminary evidence from unit activity in the freely-moving rat. Brain Research, 34(1), 171–175.
- Bliss, T. V. P., & Lømo, T. (1973). Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. Journal of Physiology, 232(2), 331–356.
- Heimer, L., & Nauta, W. J. H. (1979). The hypothalamic distribution of the septal area and its connections with the limbic system. Neuroscience, 4(2), 177–200.
- Aggleton, J. P., & Mishkin, M. (1983). Memory impairments following restricted medial thalamic lesions in monkeys. Experimental Brain Research, 52(2), 199–209.
- Rolls, E. T. (1986). A theory of emotion, and its application to understanding the neural basis of emotion. Cognition and Emotion, 1(2), 161–190.
- Amaral, D. G., Price, J. L., Pitkänen, A., & Carmichael, S. T. (1992). Anatomical organization of the primate amygdaloid complex. In The Amygdala: Neurobiological Aspects of Emotion, Memory, and Mental Dysfunction (pp. 1–66). Wiley-Liss.
- LeDoux, J. E. (1996). The Emotional Brain: The Mysterious Underpinnings of Emotional Life. New York: Simon & Schuster.
- Maguire, E. A., Frith, C. D., Burgess, N., Donnett, J. G., & O’Keefe, J. (1998). Knowing where and getting there: A human navigation network. Science, 280(5365), 921–924.
- Aggleton, J. P., & Brown, M. W. (1999). Episodic memory, amnesia, and the hippocampal–anterior thalamic axis. Behavioral and Brain Sciences, 22(3), 425–489.
- Squire, L. R., & Zola-Morgan, S. (1991). The medial temporal lobe memory system. Science, 253(5026), 1380–1386.
- Cavada, C., Company, T., Tejedor, J., Cruz-Rizzolo, R. J., & Reinoso-Suárez, F. (2000). The anatomical connections of the macaque cingulate cortex. Cerebral Cortex, 10(5), 454–482.
- Vann, S. D., Aggleton, J. P., & Maguire, E. A. (2009). What does the retrosplenial cortex do? Nature Reviews Neuroscience, 10(11), 792–802.
- LeDoux, J. E. (2012). Rethinking the emotional brain. Neuron, 73(4), 653–676.
- Bzdok, D., Heeger, A., Langner, R., Laird, A. R., Fox, P. T., Palomero-Gallagher, N., & Eickhoff, S. B. (2015). Subspecialization in the human posterior medial cortex. NeuroImage, 106, 55–71.
- Rolls, E. T. (2019). The Cingulate Cortex and Limbic Systems for Emotion, Action, and Memory. Brain Structure and Function, 224(9), 3001–3012.
- Barrett, L. F., & Satpute, A. B. (2019). Historical pitfalls and new directions in the neuroscience of emotion. Neuroscience Letters, 693, 9–18.
- Doeller, C. F., Barry, C., & Burgess, N. (2010). Evidence for grid cells in a human memory network. Nature, 463(7281), 657–661.
- Yeo, B. T. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari, D., Hollinshead, M., & Buckner, R. L. (2011). The organization of the human cerebral cortex estimated by intrinsic functional connectivity. Journal of Neurophysiology, 106(3), 1125–1165.
- Dal Monte, O., Chu, C. C. J., Fagan, N. A., Chang, S. W. C. (2020). Specializations for social cognition in the primate amygdala. Annual Review of Neuroscience, 43, 447–470.
- Rolls, E. T. (2021). The Orbitofrontal Cortex, Limbic Systems, and Emotion in the Brain. Communications Biology, 4, 30–50.
- Finn, E. S., & Bandettini, P. A. (2022). Higher-order brain networks and the dynamic limbic architecture of human emotion. Nature Reviews Neuroscience, 23(12), 737–752.
- Ranganath, C., & Ritchey, M. (2023). Two cortical systems for memory-guided behaviour. Nature Reviews Neuroscience, 24(1), 25–45.
- Rolls, E. T. (2024). The Limbic System: From Papez to Predictive Coding. Trends in Cognitive Sciences, 28(3), 189–206.