Influenza A virüsü
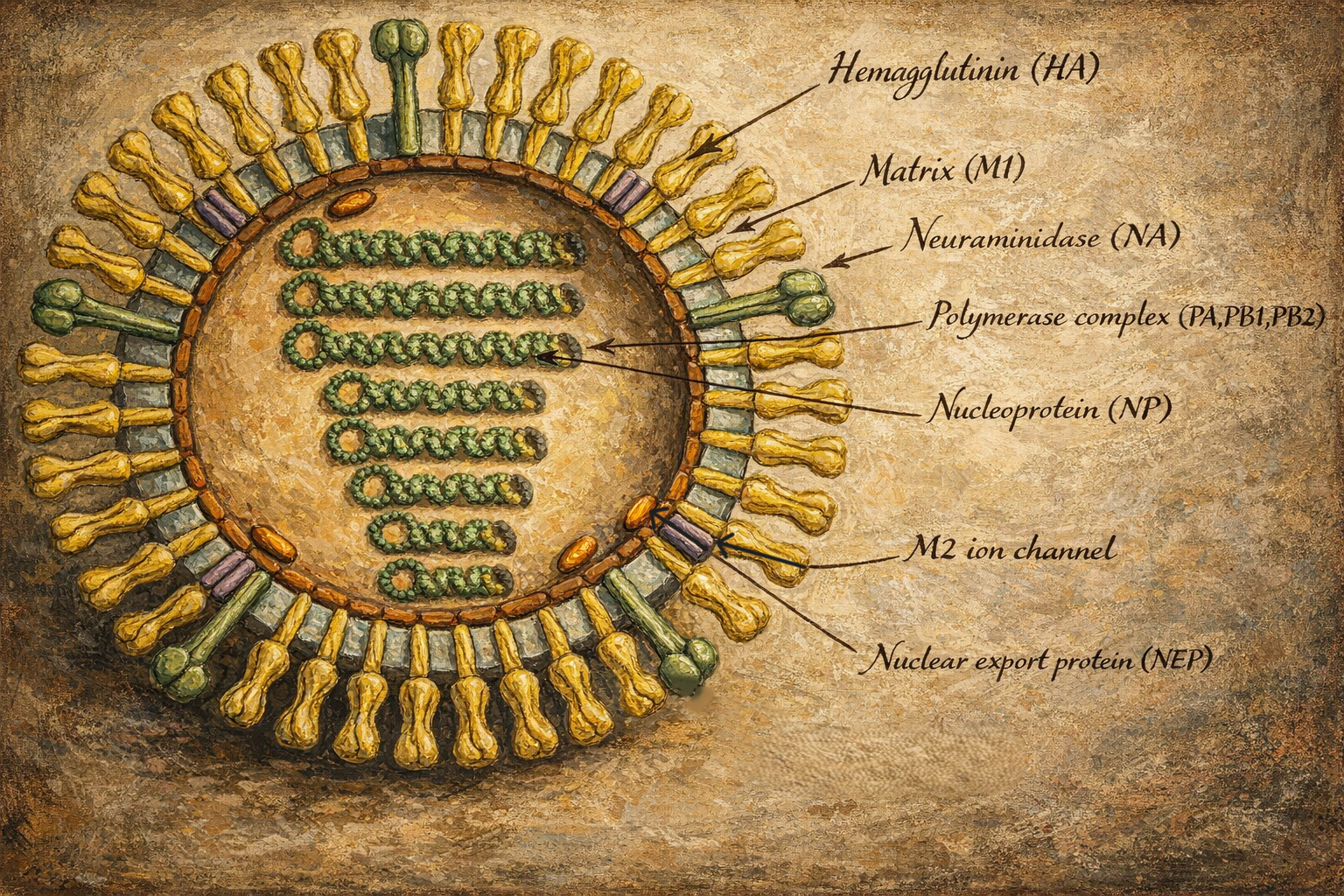
“İnfluenza” sözcüğü, İtalyanca influenza kelimesinden türemiştir ve tarihsel olarak “etki, tesir, nüfuz” anlam alanına dayanır. Orta Çağ ve erken modern dönemde salgın hastalıkların gök cisimlerinin ya da mevsimsel “havaların etkisi” ile ortaya çıktığı fikri yaygın olduğundan, terim başlangıçta hastalığın nedeni olarak varsayılan dışsal etkilenimi ima eder. Zaman içinde kavram, klinik olarak ani başlangıçlı ateş, miyalji, … Devamını oku