
Etimoloji
Lokelma ticari adı, hiperkalemiyi tedavi etmek için tasarlanmış sodyum zirkonyum siklosilikat için özel olarak tasarlanmıştır. AstraZeneca, ismin türetilmesini kamuoyuna açıklamamış olsa da, dilbilimsel ve farmasötik adlandırma;
Anlamsal Bileşenler ve Çıkarım:
“Lo-” öneki, metaforik olarak ilacın etki mekanizmasına karşılık gelen “kilitleme” kavramını çağrıştırabilir. Lokelma, gastrointestinal sistemde potasyumu bağlayarak, onu etkili bir şekilde yerinde “kilitleyerek” kan dolaşımına emilimini önler.
“-kelma” eki, muhtemelen potasyum seviyeleriyle ilgili tıbbi terminolojide kök salmış olan “kalemi” terimine gönderme yapıyor (yüksek potasyum için “hiperkalemi”de görüldüğü gibi). Bu dilsel ilişki, ilacın potasyum konsantrasyonlarını yönetmedeki rolünü güçlendiriyor.
- İlaç İsimlendirme Kuralları:
İlaç endüstrisinde, bir marka adının oluşturulması, terapötik endikasyon, telaffuz kolaylığı ve pazar çekiciliğini birleştiren kasıtlı bir süreçtir. Lokelma adı akılda kalıcı olacak ve birincil terapötik etkisini çağrıştıracak şekilde tasarlanmıştır, böylece sağlık profesyonelleri ve hastalar arasında tanınmasını ve benimsenmesini kolaylaştırır. - Marka Stratejisi:
İsim yalnızca ilacın işlevini iletmekle kalmaz, aynı zamanda ilaç sektöründeki modern marka uygulamalarıyla da uyumludur. Ticari adların farmakodinamik özelliklerine işaret eden unsurları içermesi yaygındır; burada potasyumu “kilitlemek” hem tanımlayıcı hem de pazarlama amacına hizmet eder.
Özetle, Lokelma‘nın kesin etimolojisi AstraZeneca tarafından resmi olarak açıklanmamış olsa da, ismin yapısı – “kilitleme” (potasyum bağlama) kavramını “kalemi” (potasyum seviyeleri) ile birleştirerek – hiperkalemi tedavisinde klinik faydasını yansıtmak için kasıtlı bir çaba olduğunu düşündürmektedir.
Marka: AstraZeneca
Etkin Madde: Sodyum zirkonyum siklosilikat
Farmakolojik Sınıf: Potasyum bağlayıcı
Formülasyon: Oral toz
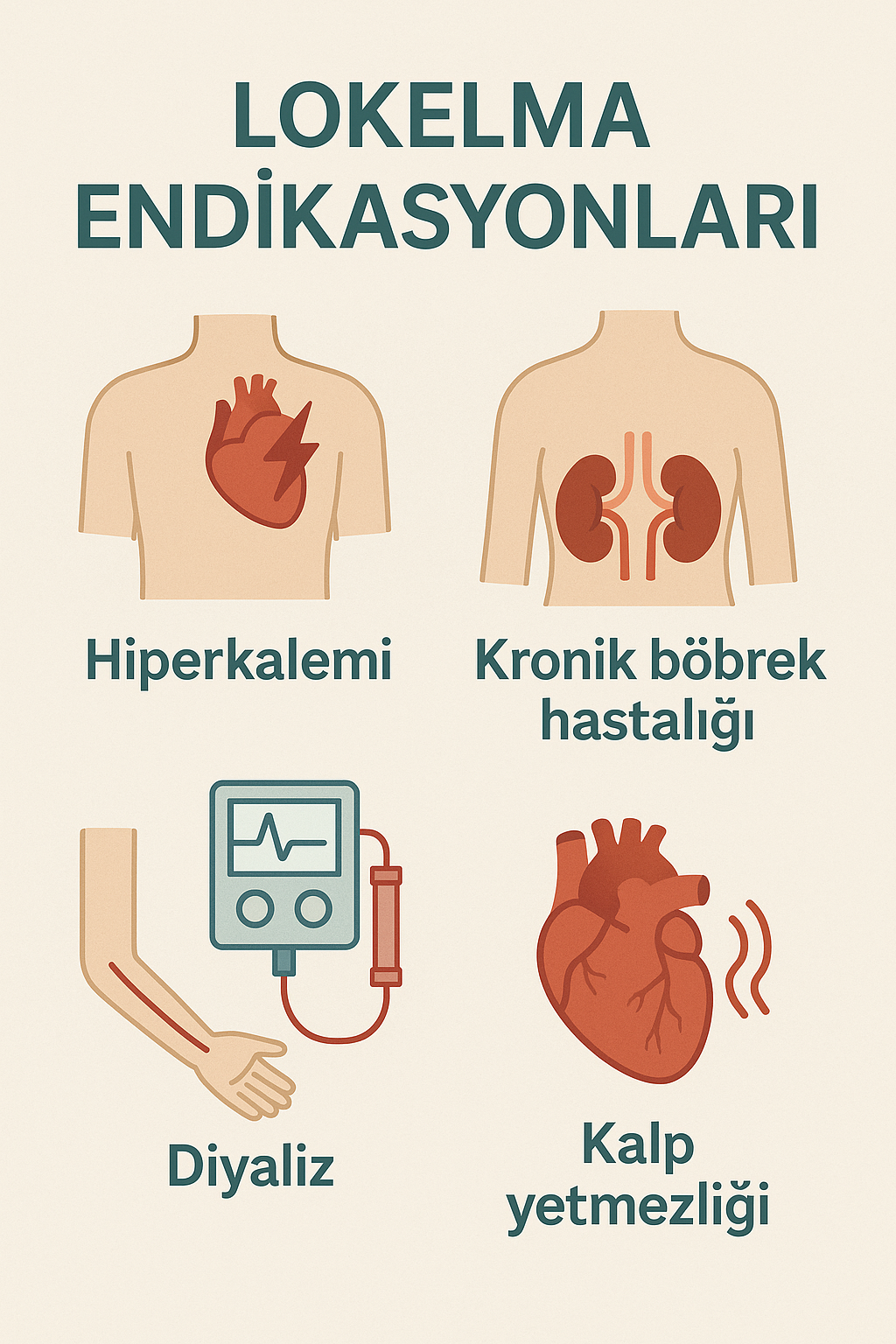
Endikasyon:
Lokelma, yetişkin hastalarda serum potasyum seviyelerinin yükselmesiyle karakterize bir durum olan hiperkaleminin yönetimi için endikedir. Hiperkalemi, böbrek yetmezliği, kalp yetmezliği veya potasyum homeostazını değiştiren ilaçların kullanımı dahil olmak üzere çeşitli etiyolojilerden kaynaklanabilir.
Dozaj ve Uygulama:
- Dozaj: Önerilen doz genellikle oral yoldan uygulanan 5 gram veya 10 gramdır. Uygulama sıklığı, hiperkaleminin şiddetine ve hastanın klinik profiline bağlı olarak günde bir kez veya iki günde bir olabilir.
- Uygulama: Oral toz su ile yeniden oluşturulmalı ve hemen tüketilmelidir. Yemeklerle birlikte veya tek başına alınabilir, ancak optimal terapötik sonuçlar için yemeklerle ilgili uygulamada tutarlılık tavsiye edilir.
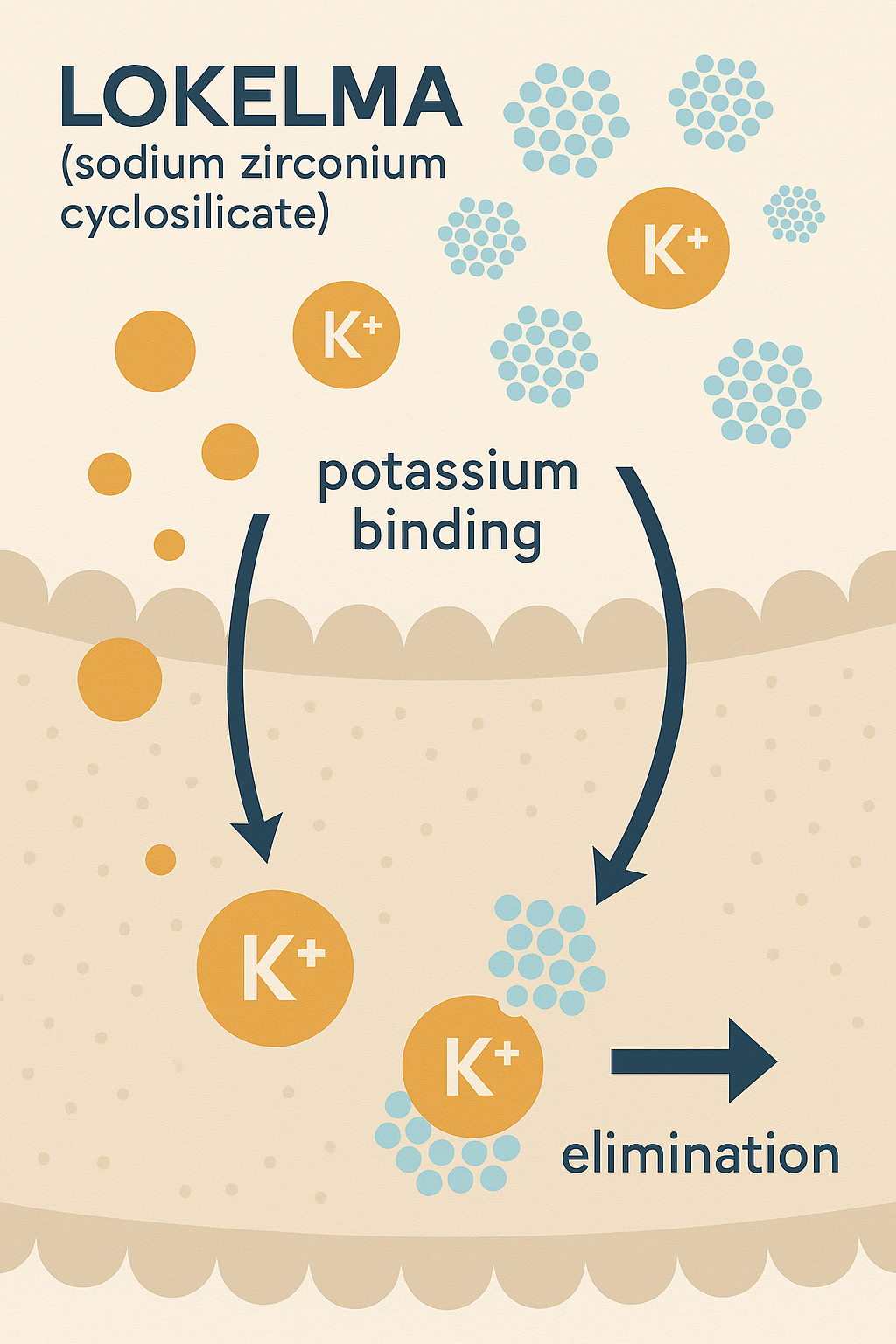
Etki Mekanizması:
Lokelma, terapötik etkisini gastrointestinal sistemdeki potasyum iyonlarını seçici olarak bağlayarak gösterir. Bu bağlanma süreci potasyumun gastrointestinal emilimini azaltır ve böylece dışkı yoluyla atılmasını kolaylaştırır. Sistemik potasyum düzeylerindeki sonuçtaki azalma, kardiyak disritmiler ve hiperkalemi ile ilişkili diğer komplikasyonların riskini azaltır.
Klinik Kullanım:
Serum potasyumunu düşürmedeki etkinliği nedeniyle Lokelma, hiperkaleminin yönetiminde kullanılır. Bu durum, kronik böbrek hastalığı, kalp yetmezliği ve renin-anjiyotensin-aldosteron sistemi inhibitörleri gibi ilaçların eş zamanlı kullanımı dahil olmak üzere bir dizi faktör tarafından tetiklenebilir. Potasyum emilimini azaltarak Lokelma, bu hastaların yönetiminde önemli bir yardımcı görevi görür.
Olumsuz Etkiler:
- Yaygın Olumsuz Etkiler: Gastrointestinal rahatsızlıklar en sık bildirilen yan etkilerdir ve ishal, mide bulantısı, kusma, kabızlık ve karın ağrısını içerir.
- Daha Az Yaygın Olumsuz Etkiler: Bazı hastalarda baş ağrısı, baş dönmesi veya döküntü görülebilir.
Uyarılar ve Önlemler:
- Aşırı duyarlılık: Lokelma, sodyum zirkonyum siklosilikata karşı bilinen bir alerjisi veya aşırı duyarlılığı olan hastalarda kontrendikedir.
- Belirli Popülasyonlarda Dikkat: Böbrek yetmezliği, kalp yetmezliği veya dehidratasyonu olan hastalarda dikkatli kullanın. Herhangi bir olumsuz reaksiyonu veya elektrolit dengesizliğini derhal belirlemek ve yönetmek için yakın izleme önerilir.
- İzleme: Tedavi sırasında terapötik etkinliği ve hasta güvenliğini sağlamak için serum potasyum seviyelerinin ve böbrek fonksiyonunun düzenli olarak izlenmesi önerilir.
Keşif
Erken Kavramsallaştırma ve Klinik Öncesi Geliştirme (2000’ler – 2010’ların Başları):
- İhtiyacın Belirlenmesi: 2000’lerin başlarında, klinik gözlemler ve araştırmalar, hiperkalemiyi tedavi etmede mevcut potasyum bağlayıcıların (sodyum polistiren sülfonat gibi) sınırlamalarını vurguladı. Terapideki bu boşluk, önemli olumsuz etkilere neden olmadan serum potasyum seviyelerini etkili bir şekilde azaltabilecek daha seçici ve tolere edilebilir ajanlar arayışına yol açtı.
- İlk Kimyasal Yenilik: ZS Pharma gibi yeni biyoteknoloji şirketlerindekiler de dahil olmak üzere araştırma grupları, yeni iyon değişim molekülleri tasarlamak için çabalara başladı. Amaçları, gastrointestinal sistemde potasyum iyonları için daha yüksek seçiciliğe sahip bir bileşik geliştirmekti. 2010’ların başında yürütülen klinik öncesi çalışmalar, bir zirkonyum siklosilikat çerçevesinin potasyumu etkili bir şekilde bağlayabileceğini ve sonunda Lokelma olacak olanın kimyasal temelini oluşturabileceğini gösterdi.
Translasyonel Araştırma ve Erken Klinik Denemeler (2012–2014):
- Klinik Öncesi Etkinlik ve Güvenlik Çalışmaları: In vitro ve hayvan modeli çalışmaları, sodyum zirkonyum siklosilikatın seçiciliği ve bağlanma kapasitesi hakkında kritik veriler sağladı. Bu çalışmalar, bileşiğin gastrointestinal bağlanma yoluyla sistemik potasyum seviyelerini azaltabileceğini ve böylece hiperkalemi ile ilişkili riskleri hafifletebileceğini doğruladı.
- Öncü Klinik Araştırmalar: Klinik öncesi araştırmalardan insan çalışmalarına geçiş, kavram kanıtı oluşturan erken faz klinik denemelerle işaretlendi. Dr. Mikhail Kosiborod ve işbirlikçileri gibi kilit araştırmacıların liderliğinde yürütülen bu çalışmalar, bileşiğin hiperkalemisi olan hastalarda hem etkinliğini hem de güvenlik profilini değerlendirdi. Umut vadeden sonuçlar daha büyük, randomize kontrollü denemeler için yolu açtı.
Büyük Ölçekli Klinik Değerlendirme ve Düzenleyici Önemli Noktalar (2014–2018):
- HARMONIZE Denemesi (2014): En önemli önemli noktalardan biri, ilacın tedaviden sonraki 28 gün içinde serum potasyum seviyelerini düşürme yeteneğine dair sağlam kanıtlar sağlayan çok merkezli, randomize klinik bir deneme olan HARMONIZE denemesiydi. Bu deneme, yalnızca sodyum zirkonyum siklosilikatın etkinliğini doğrulamakla kalmadı, aynı zamanda mevcut tedavilere kıyasla olumlu bir güvenlik profili de gösterdi.
(Ana Katkıda Bulunan: Dr. Kosiborod ve ekibi bu çalışmada önemli bir rol oynadı.) - Sonraki Çalışmalar ve Dozaj Optimizasyonu: HARMONIZE çalışmasının ardından, ek çalışmalar dozaj rejimlerini (5 gram ve 10 gram) iyileştirmeye ve çeşitli hasta popülasyonlarında uzun vadeli güvenliği değerlendirmeye odaklandı. Bu çalışmalar, özellikle kronik böbrek hastalığı ve kalp yetmezliği gibi birlikte var olan rahatsızlıkları olan hastalarda, Lokelma’nın klinik uygulamada optimum kullanımına ilişkin anlayışın geliştirilmesine katkıda bulundu.
- Düzenleyici Onay (2018): Bu klinik araştırmalardan elde edilen kanıtların birikimi, 2018 yılında ABD Gıda ve İlaç Dairesi (FDA) gibi kurumlar tarafından düzenleyici onayla sonuçlandı. Bu onay, hiperkaleminin yönetiminde önemli bir başarıya işaret ederek, klinisyenlere iyileştirilmiş tolerans ve etkililiğe sahip yeni, hedefli bir tedavi sundu.
Onay Sonrası Gelişmeler ve Devam Eden Araştırma (2018–Günümüz):
- Klinik Uygulamaya Entegrasyon: Düzenleyici onayın ardından Lokelma, hiperkalemi için tedavi kılavuzlarına giderek daha fazla dahil edildi. Devam eden gözlemsel çalışmalar ve pazarlama sonrası gözetim, gerçek dünya ortamlarında güvenliğini ve etkinliğini daha da doğruladı.
- Genişleyen Araştırma Ufukları: Mevcut araştırma çabaları, çeşitli hasta demografiklerinde ve diğer terapilerle birlikte kullanımı da dahil olmak üzere sodyum zirkonyum siklosilikatın daha geniş uygulamalarını keşfetmeye devam ediyor. Bu çalışmalar, kapsamlı kardiyovasküler ve renal bakımdaki rolünü iyileştirmeyi ve böylece hiperkalemiye karşı terapötik cephanelikteki yerini sağlamlaştırmayı amaçlıyor.
İleri Okuma
- Kosiborod, M., Rasmussen, H. L., Lavin, P., et al. (2014). Effect of sodium zirconium cyclosilicate on potassium lowering for 28 days among outpatients with hyperkalemia: The HARMONIZE randomized clinical trial. JAMA, 312(21), 2223-2233.
- Bakris, G. L., Pitt, B., Weir, M. R., Packham, D. K., et al. (2015). Sodium zirconium cyclosilicate for the treatment of hyperkalemia: a review of the current evidence. Journal of Nephrology, 28(5), 573-581.
- Pitt, B., Weir, M. R., Ball, M., et al. (2016). Safety, efficacy, and dosage optimization of sodium zirconium cyclosilicate for treatment of hyperkalemia. The American Journal of Cardiology, 118(4), 656-662.
- Lokelma (sodium zirconium cyclosilicate) – DailyMed: https://dailymed.nlm.nih.gov/dailymed/lookup.cfm?setid=90bf8e28-748d-4e4b-a19f-9cf483370eff
- Clinical Review Report: Sodium Zirconium Cyclosilicate (Lokelma): https://www.ncbi.nlm.nih.gov/books/NBK563715/
- Pharmacoeconomic Review Report: Sodium Zirconium Cyclosilicate (Lokelma): https://www.ncbi.nlm.nih.gov/books/NBK563720/
- Highlights of Prescribing Information: Lokelma: https://www.accessdata.fda.gov/drugsatfda_docs/label/2018/207078s000lbl.pdf
- Sodium zirconium cyclosilicate – Wikipedia: https://en.wikipedia.org/wiki/Sodium_zirconium_cyclosilicate




Yorum yazabilmek için oturum açmalısınız.