Düz kasın hikayesi, antik anatomik çalışmalardan modern moleküler biyolojiye kadar yüzyılları kapsayan bilimsel anlayışın gelişen doğasının bir kanıtıdır. Görünür hareketleri ve lokomosyondaki rolü nedeniyle erken dönemde dikkat çeken iskelet kasının aksine, düz kas arka planda kalmış ve vücutta sessizce temel işlevleri yerine getirmiştir. Düz kas anlayışımızdaki dönüm noktaları, yeni teknolojiler, değişen bakış açıları ve bu dokunun gizli inceliklerinin kademeli olarak ortaya çıkarılması arasındaki büyüleyici etkileşimi ortaya koymaktadır.
Erken Dönem Gözlemler: Antik Çağ’dan Rönesans’a (Antik Yunan – 1600’ler)
Kas dokusu kavramı eski uygarlıklara, özellikle de insan ve hayvan diseksiyonları üzerine yaptığı çalışmalarda kasların anatomisini tanımlayan Galen (MS 2. yüzyıl) gibi Yunan hekimlerin yazılarına dayanmaktadır. Ancak, farklı kas dokusu türleri arasında ayrım yapmamıştır. Açıklamaları, hareketteki rolü açık ve gözlemlenebilir olan daha belirgin, çizgili kaslara odaklanmıştır.
Rönesans’a kadar, 16. yüzyılda Andreas Vesalius tarafından yapılanlar da dahil olmak üzere, daha ayrıntılı anatomik çalışmalar eski kavramlara meydan okumaya başladı. Vesalius, ayrıntılı diseksiyonlar yoluyla kas anatomisini anlamak için zemin hazırladı, ancak düz kas, iç organların duvarları içinde daha derin, daha az göze çarpan doğası nedeniyle zor kaldı.
Mikroskobun Şafağı: Leeuwenhoek ve Malpighi (1600’ler)
Mikroskobun 17. yüzyılda Antonie van Leeuwenhoek ve Marcello Malpighi gibi öncüler tarafından icat edilmesi biyolojiyi dönüştürmüştür. İtalyan bir anatomist olan Malpighi, kılcal damarların ve kan damarlarının karmaşık yapıları da dahil olmak üzere çeşitli doku ve organların yapısını incelemek için mikroskoplar kullandı. Çalışmaları, kan damarlarının duvarlarında, iskelet dokularında incelediği çizgili kaslara benzemeyen farklı bir kas türünün varlığına işaret ediyordu.
Leeuwenhoek, mikroskobik gözlemlerinde çeşitli dokulardaki kas liflerini tanımladı. Kas liflerinin görünümündeki farklılıklara dikkat çekti, ancak düz ve çizgili kaslar arasında temel bir ayrım olduğu henüz net değildi.
Düz Kasın Benzersiz Olarak Tanınması: 19. Yüzyıl Keşifleri
Bilim insanları hücresel yapıları daha ayrıntılı incelemek için gelişmiş mikroskopi tekniklerini kullandıkça, 19. yüzyıl histoloji ve doku sınıflandırmasında önemli ilerlemelere sahne oldu. Friedrich Henle ve Albert von Kölliker gibi Alman bilim insanları, düz kas da dahil olmak üzere dokuların diğer kas türlerinden farklı olarak sınıflandırılmasında kritik roller oynadı.
Anatomi ve histolojiye yaptığı katkılarla tanınan Henle, kan damarlarının duvarlarında düz kasın varlığını tanımladı ve damar tonusunun düzenlenmesindeki rolünü belirledi. Kölliker’in çalışmaları kas dokularının anlaşılmasını daha da geliştirdi. Düz kası, iğ şeklindeki hücrelerine ve iskelet ve kalp kaslarının ayırt edici özelliği olan görünür çizgilerin eksikliğine dayanarak çizgili kastan ayırdı. Bu, düz kası kendi yapısı ve işlevi olan benzersiz bir kas dokusu türü olarak belirlediği için çok önemli bir dönüm noktasıydı.
İşlevi Anlamak: 20. Yüzyılın Başlarında Fizyolojiye İlişkin Anlayışlar
Düz kasın benzersiz bir doku olarak tanımlanmasıyla birlikte, odak noktası işlevini anlamaya doğru kaymıştır. 20. yüzyılın başları, özellikle istemsiz fonksiyonları kontrol eden otonom sinir sistemi ile ilişkili olarak düz kas fizyolojisi üzerine yapılan çalışmalarla damgasını vurmuştur.
Amerikalı bir fizyolog olan Walter Cannon bu alanda önemli katkılarda bulunmuştur. “Savaş ya da kaç” tepkisi üzerine yaptığı araştırmalar, otonom sinir sisteminin strese yanıt olarak düz kasların kasılmasını ve gevşemesini nasıl kontrol ettiğini, kan akışı ve sindirim gibi süreçleri nasıl etkilediğini ortaya koymuştur. Bu çalışma, düz kasın hem iç hem de dış uyaranlara yanıt vererek homeostazın korunmasındaki rolünü vurguladı.
“Tonus” veya tonik kasılma kavramı bu dönemde ortaya çıkmış ve düz kasın uzun süreler boyunca kısmi kasılma durumunu sürdürme yeteneğini vurgulamıştır. Bu özellik, kan basıncını düzenlemesi gereken kan damarları ve yiyecekleri sindirim sistemi boyunca hareket ettirmek için sürekli kasılmalar gerektiren gastrointestinal sistem gibi organların işleyişi için çok önemlidir.
Biyokimyasal Atılımlar: Aktin ve Miyozinin Keşfi (1940’lar-1960’lar)
- yüzyılın ortaları, kas kasılmasının altında yatan moleküler mekanizmaların anlaşılmasında çığır açtı. İskelet kasındaki aktin ve miyozinin keşfine dayanarak, araştırmacılar bu proteinlerin düz kasta nasıl işlev gördüğünü keşfetmeye başladılar.
1940’larda ve 1950’lerde yapılan çalışmalar, düz kasın da kasılma için aktin ve miyozine dayandığını, ancak önemli bir farkla: bu proteinlerin organizasyonunun daha az düzenli olduğunu ve düz kasın birden fazla yönde kuvvet üretmesine izin verdiğini ortaya koydu. Çizgili kasın düzenli sarkomerlerinin aksine, düz kasın aktin ve miyozin filamentleri çapraz bir şekilde düzenlenerek kasın geniş bir uzunluk aralığında kasılmasını ve gerginliğini korumasını sağlar.
Bu dönemde kalsiyumun kas kasılmasındaki rolü de keşfedilmiştir. Kalsiyum iyonunun (Ca²⁺) hem çizgili hem de düz kaslarda kasılmaların tetiklenmesinde çok önemli olduğu bulunmuştur. Bununla birlikte, düz kasta kalsiyum, çizgili kasta görülen troponin-tropomiyozin sisteminden ziyade, protein kalmodulin ve miyozin hafif zincir kinaz (MLCK) adı verilen bir enzimi içeren farklı bir mekanizma aracılığıyla işlev görür. Bu keşifler, düz kas işlevini yöneten benzersiz düzenleyici yolların anlaşılması için temel oluşturdu.
Modern Dönem: Moleküler ve Klinik Anlayışlar (1970’ler-Günümüz)
1970’ler ve sonrası moleküler biyoloji çağına damgasını vurdu ve düz kasları kontrol eden sinyal yolları ve düzenleyici mekanizmalar hakkında daha derin bilgiler sağladı. Araştırmacılar, düz kas hücrelerinde aktin ve miyozin arasındaki etkileşimi modüle eden kaldesmon ve kalponin gibi proteinlerin rollerini ortaya çıkardı. Bu keşifler, çeşitli fizyolojik taleplere yanıt olarak düz kas kasılmasının nasıl ince ayarlandığına dair daha incelikli bir anlayış sağladı.
Bu dönemde bilim insanları düz kas fonksiyonunu hedef alan farmakolojik ajanlar da geliştirdi. Düz kas hücrelerine kalsiyum akışını etkileyen kalsiyum kanal blokerleri gibi ilaçlar, kan damarlarındaki düz kasları gevşeterek hipertansiyon ve anjin gibi durumların tedavisinde önemli araçlar haline geldi. Temel araştırmalar ve klinik uygulamalar arasındaki bu bağlantı, düz kas anlayışının tıptaki önemini vurgulamıştır.
Modern çağda konfokal mikroskopi ve elektron mikroskobu gibi görüntüleme tekniklerindeki ilerlemeler, bilim insanlarının düz kas hücrelerindeki aktin, miyozin ve diğer proteinlerin karmaşık etkileşimlerini görselleştirmelerine olanak sağladı. Ayrıca, genetik ve moleküler çalışmalar, akciğerlerin hava yollarından mesaneye kadar farklı dokulardaki düz kas hücrelerinin çeşitliliğini ortaya çıkarmış ve her biri kendi özel rolüne adapte olmuştur.
Düz kas çalışmaları, eski anatomistlerin spekülatif gözlemlerinden günümüzün karmaşık moleküler modellerine kadar uzun bir yol kat etmiştir. Düz kas tarihindeki her kilometre taşı, benzersiz yapısal organizasyonundan işlevini kontrol eden biyokimyasal yollara kadar yeni anlayış katmanları getirmiştir. Bu yolculuk sadece vücudumuzun nasıl çalıştığına dair bilgilerimizi zenginleştirmekle kalmamış, aynı zamanda kardiyovasküler tıp, gastroenteroloji ve solunum sağlığı gibi alanları etkilemeye devam eden pratik uygulamalara da yol açmıştır.
Düz kasın hikayesi, bariz olanın ötesine bakmanın önemini vurgulamaktadır. İskelet kasının güçlü kasılmaları bir zamanlar ilk anatomistlerin dikkatini çekmiş olsa da, kan akışından sindirime kadar her şeyi düzenleyerek yaşamı sürdürmek için gerekli olduğu kanıtlanmış olan düz kasın daha ince, sabit kasılmalarıdır. Ve bilim ilerlemeye devam ettikçe, düz kas, gizli karmaşıklıkları hala çözülmesi gereken yeni gizemler sunan hayati bir çalışma konusu olmaya devam ediyor.


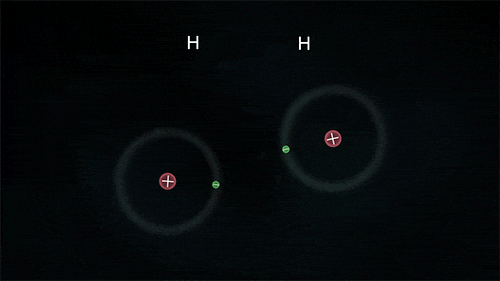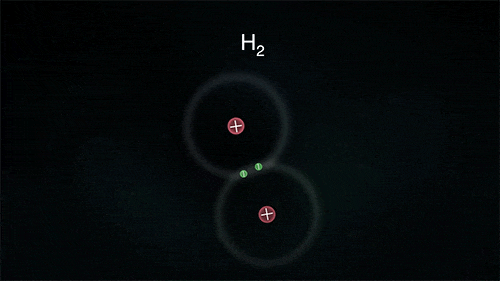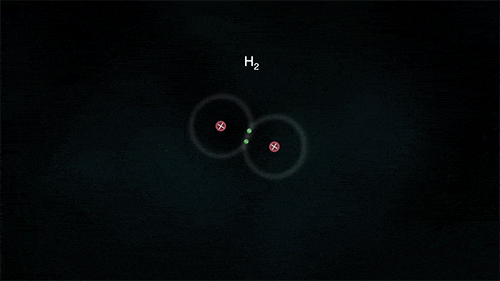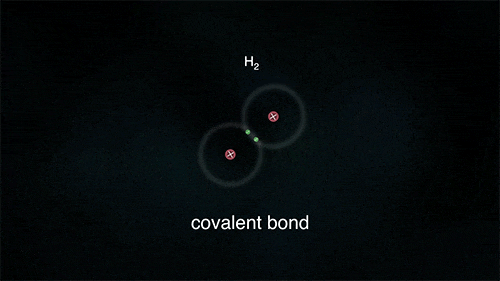
 tarafından adlandırılan,kovalent olmayan zayıf çekim kuvvetidir.
tarafından adlandırılan,kovalent olmayan zayıf çekim kuvvetidir.




 uğu fizyolojik mekanizma, bir organın değişen kan basıncına karşı aldığı önlemdir.
uğu fizyolojik mekanizma, bir organın değişen kan basıncına karşı aldığı önlemdir.