İç kulağın “tüy hücreleri”nin hikâyesi, ince cam mercekler altında beliren belirsiz çizgilerle başlar ve günümüzde, tekil iyon kanallarının atomik benzetimlerine, kaybolan işitmenin geri kazanılmasını hedefleyen klinik denemelere kadar uzanır. Bu, aynı zamanda bilimin nasıl ilerlediğinin bir örneğidir: sezgisel gözlemler, yavaş yavaş daha keskin aletlerle doğrulanır; yanlış hipotezler, daha iyi olanlarla yer değiştirir; ve kavrayış, anatomiden fiziğe, fizikten moleküle doğru katman katman derinleşir.
1) Erken İzler: Ön-Odiyoloji Çağı (17.–18. yüzyıllar)
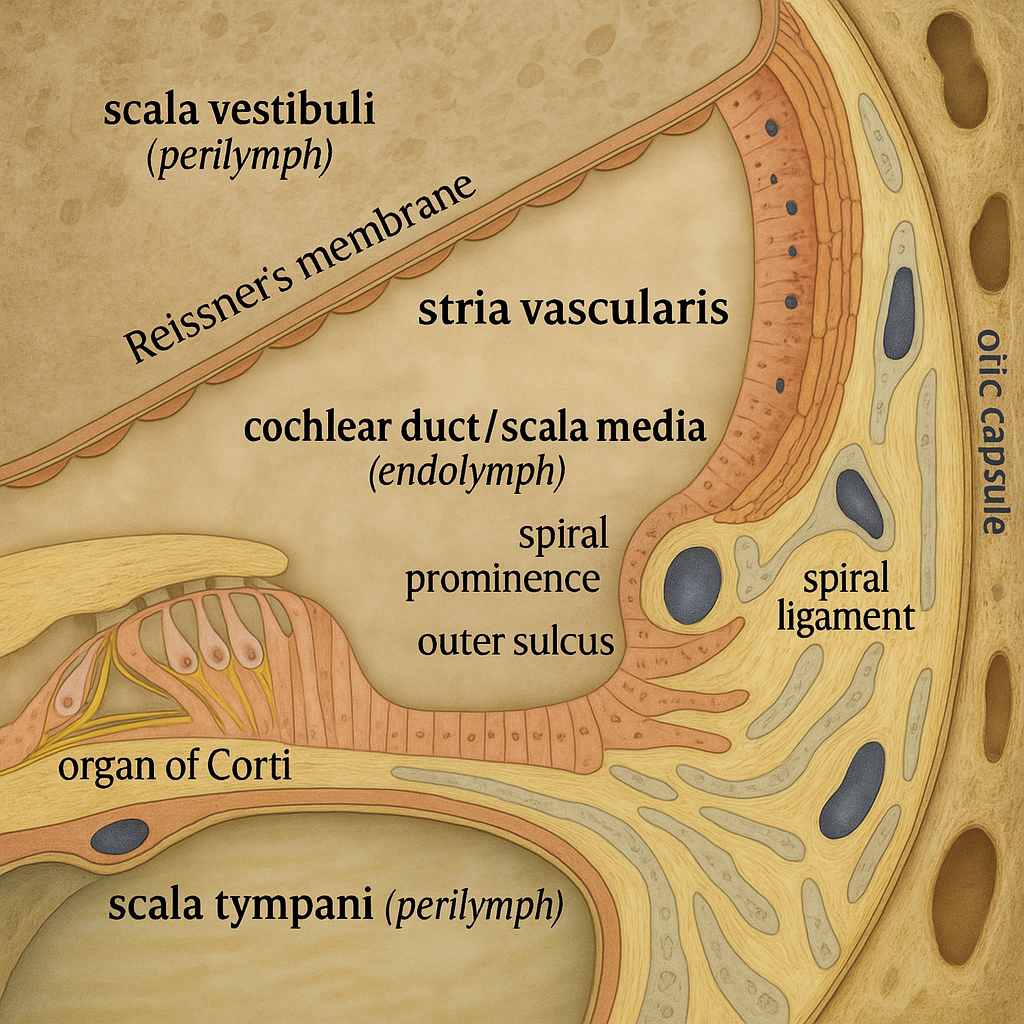
Hikâye, 17. yüzyılın ikinci yarısında, mikroskobinin doğum sancılarıyla başlar. Marcello Malpighi, ince kesitler ve ham optiklerle memeli iç kulağının kıvrımlarını betimler; onun notlarında, bugün bizim “stereosilya” diyeceğimiz saç benzeri çıkıntılara doğrudan göndermeler beklemek aşırı olur, fakat labirentin dokusal ayrıntılarının ilk sistematik tasvirlerine giden yolu açtığı tartışmasızdır. Birkaç on yıl sonra Antonio Valsalva, temporal kemiğin ve labirentin topografyasını klinik gözlemlerle birleştirir; Domenico Cotugno ise 18. yüzyılın ortasında labirentin sıvılarını –sonradan “endolenf” ve “perilenf” olarak adlandırılacak ayırtları– dikkat çekici bir berraklıkla tasvir eder. Henüz “tüy hücresi” terimi yoktur; fakat sahne kurulmuştur: katı kemik kapsül içinde akışkan bir dünya.
2) Mikroskobun Netleşmesi ve Organın Adını Bulması (19. yüzyılın ortası)
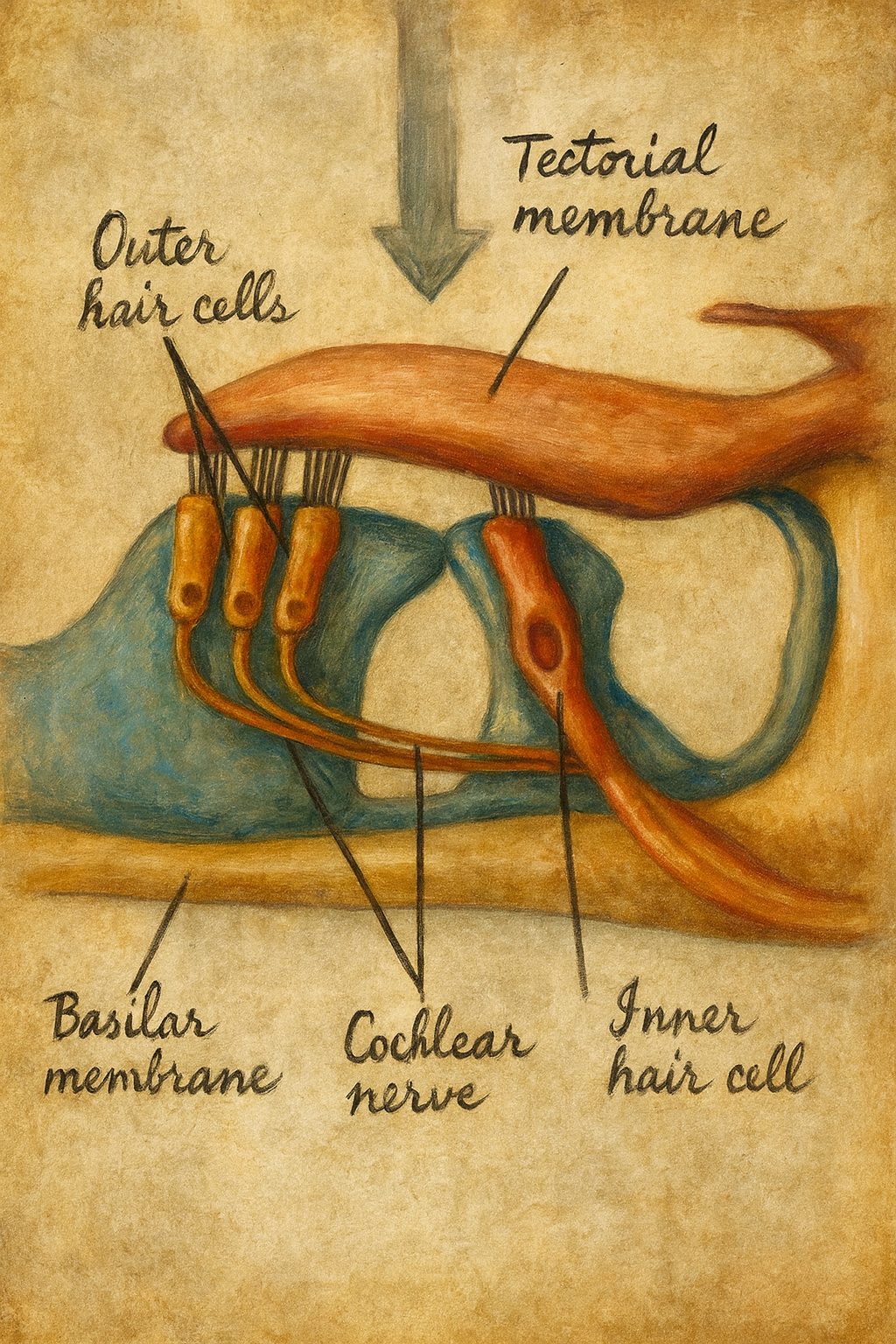
Modern anlamda keşif, 1851’de Alfonso Corti’nin tarihî çalışmasıyla görünür hâle gelir. Corti, memeli kokleasında baziler membran üzerinde uzanan sinir-epitelyal şeridi ayrıntılarıyla tanımlar; bugün “Corti organı” dediğimiz bu yapı, iç (tek sıra) ve dış (üç sıra) hücre dizilerinin anatominin omurgası olduğunu ortaya koyar. Aynı dönemde Otto Deiters destek hücrelerini, Victor Hensen ve Friedrich Matthias Claudius epitelin eşlikçi hücre tiplerini betimler; Albert von Kölliker ve diğer histologlar, kemik labirentin içinde benzersiz bir sensör-mekanik organizasyonun ipuçlarını resmeder. Burada “keşif” artık yalnız bir görme olayı değildir: farklı hücre tiplerinin düzeni ve muntazam dizilişi, bir işlevi ima eder.
3) “Nasıl Duyuyoruz?”: Kuramsal Sıçrama ve Vestibüler Kavrayış (19. yüzyıl sonu)
Hermann von Helmholtz, 1863’te yayımladığı ses duyumları kuramıyla, kokleanın frekanslara göre yerel olarak ayarlı bir rezonatörler dizisi olabileceğini ileri sürer. Bu fikir, henüz doğrudan ölçülemeyen bir şeyi –frekans seçiciliği– anatomik düzen ile uzlaştırır. Öte yandan Ernst Ewald’ın 1890’larda yarım daire kanalları üzerine yaptığı titiz deneyler, vestibüler sistemin yön-seçici ve dinamik yanıtlarını nicel hâle getirir; Robert Bárány’nın klinik denemeleri, termal uyarımla vestibüler reflekslerin ortaya çıkarılabileceğini gösterir ve 1914’te Nobel Ödülü’yle taçlanır. Bir yanda işitmenin mekaniği, öte yanda dengenin refleks fiziolojisi… her ikisi de aynı hücresel ailenin –tüy hücrelerinin– omuzlarında yükselir.
4) Optikten Elektrofizyolojiye: Dalgaların İzini Sürmek (20. yüzyıl başı–ortası)
Georg von Békésy, 1930’lardan 1950’lere uzanan bir dizi parlak deneyle, baziler membranda seyahat eden dalganın varlığını doğrudan gösterir: belirli bir frekans, kokleanın belli bir yerinde tepe yapar. Helmholtz’un sezgisi ölçüye dönüşür; 1961’de Nobel Ödülü gelir. Ancak Békésy’nin preparatları çoğu kez postmortem ve pasif dokulardır; bu, kokleanın yalnızca pasif bir filtre olmadığı düşüncesini filizlendirir. Thomas Gold, 1948’de “aktif” bir süreç öngörür; o gün için cesur bir hipotez olan bu fikir, ileride dev bir kapıyı aralayacaktır. Aynı yıllarda Rasmussen, beyin sapından kokleaya inen olivokoklear lifleri betimler; periferdeki kazanç kontrolünün merkezi sinir sistemi tarafından modüle edildiği anlaşılır.
5) Görünmeyeni Görmek: Elektron Mikroskopisi ve Tüy Hücresi Morfogenezi (1950–1970)
Ernst Ruska’nın açtığı yoldan ilerleyen elektron mikroskopisi, 1950’lerden itibaren iç kulağın ultra-yapısını görünür kılar. Torsten Wersäll, vestibüler epitelyumda Tip I ve Tip II tüy hücrelerini ayırt eder; Engström ve meslektaşları koklear stereosilya demetlerinin kademe düzenini, apikal “uç-bağlantıları”nın (tip links) varlığını ve sinaptik bantların örgüsünü tasvir eder. Artık “saç benzeri çıkıntılar” yalnızca mecaz değildir; aktin demetlerinin düzeni, kinosilyumun gelişimdeki kılavuz rolü ve retiküler laminanın mekanik bütünlüğü, yüksek çözünürlüklü görüntülerle somutlaşır.
6) Sesin Elektriğe Dönüştüğü An: Mekanotransdüksiyonun Doğrudan Kaydı (1970–1980)
Cam pipetler ve yüksek dirençli amplifikatörler, tüy hücrelerinin elektriksel dilini çözmeyi mümkün kılar. Robert Fettiplace ve çalışma arkadaşları, sürüngen papillasında tek tüy hücresinden akım kaydeder; David Corey ve A. James Hudspeth, kurbağa sakrusal organında mekanotransdüksiyon (MET) akımını doğrudan ölçer. Ian Russell ve meslektaşları, tüy demetlerine uygulanan en küçük sapmaların, milisaniyeler içinde iyon akımlarına dönüştüğünü gösterir. Aynı dönemde David Kemp, dış kulak yolundan otoakustik emisyonlar kaydeder; iç kulağın bir “mikrohoparlör” gibi de davranabildiği –yani aktif bir amplifikatöre sahip olduğu– şaşırtıcı biçimde doğrulanır.
7) Koklear Amplifikatörün Motoru: OHC Elektromotilitesi ve Prestin (1980–2000)
1985’te William Brownell, memeli dış tüy hücrelerinin membran potansiyelindeki küçük değişimlerle boylarını elektromekanik olarak anında değiştirdiğini keşfeder. Gold’un “aktif süreç” sezgisi, artık hücresel bir mekanizmaya kavuşur. Takip eden on yıllarda bu fenomenin moleküler temeli netleşir: OHC membranına gömülü bir anyon taşıyıcı ailesi üyesi olan prestin (SLC26A5), gerilimle konformasyon değiştirerek hücrenin boyunu değiştirir; prestinsiz fareler, keskin frekans ayarını ve hassasiyeti kaybeder. Koklear amplifikatörün bedeni dış tüy hücreleri, ruhu prestindir.
8) Uç-Bağlantılar, MET Kanalı ve Şeritli Sinaps: Moleküllerin Adını Koymak (1980–2015)
1984’te Andrew Pickles ve meslektaşları, komşu stereosilyaların uçlarını birleştiren “uç-bağlantıları” gösterir. 2000’lerin başında cadherin-23 ve protocadherin-15’in bu bağlantıların ana bileşenleri olduğu anlaşılır; böylece mekanik gerilimin kanala nasıl iletilebileceğine dair bir moleküler köprü kurulur. 2010’lara gelindiğinde, TMC1/TMC2 proteinleri vertebrat tüy hücrelerindeki MET kanalının gözenek oluşturucu adayları olarak öne çıkar; model organizmalar ve insan mutasyonları, bu adaylığı kuvvetlendirir. Aynı zamanda iç tüy hücrelerinin şeritli sinapslarında vezikül trafiğinin özgül düzenleyicileri –otoferlin gibi– belirlenir; afferent iletim kapasitesinin sürekliliği, sinaptik bant mimarisiyle ilişkilendirilir.
9) Rejenerasyon, Sinaptopati ve Genetik Çağ (1988–günümüz)
1988’de Edwin Rubel, Douglas Cotanche ve diğerleri, kuş kokleasında hasarlı tüy hücrelerinin destek hücrelerinden kaynaklanarak yenilenebildiğini gösterir; memelilerde ise bu kapasite şaşırtıcı biçimde sınırlıdır. 1990’lar ve 2000’lerde, kalıtsal işitme kayıplarını kodlayan onlarca gen –MYO7A, CDH23, PCDH15, KCNQ4, DIAPH1 ve daha niceleri– haritalanır; klinik fenotipler moleküler patikalarla eşleşir. 2009’dan itibaren “koklear sinaptopati” kavramı, eşikler normal iken gürültüde konuşma ayrımının bozulabileceğini, yani sinaptik bağlantıların sessizce zayıflayabileceğini gösterir. Böylece işitme kaybının yalnız “hücre ölümü”nden ibaret olmadığı, sinaptik ve nöral düzeylerde de başlayabileceği anlaşılır.
Bu arada klinik sahada bir başka devrim zaten yola çıkmıştır: 1960’lardan itibaren William House’un öncülük ettiği koklear implantlar, 1970’lerin sonlarında Graeme Clark’ın çok kanallı tasarımıyla olgunlaşır. Tüy hücreleri hasar gördüğünde, cihaz sinir liflerini doğrudan uyararak işitsel duyumu geri kazandırır; binlerce çocuğun dil edinimi ve eğitimi, bu teknoloji sayesinde kökten değişir.
10) Güncel Ufuklar: Tercüme Edilen Genler, Çözülen Yapılar, İncelen Duyumlar (2015–2025)
Son on yılda, üç doğrultu belirginleşir:
- Gen ve hücre tedavileri: Özellikle bialelik OTOF mutasyonları gibi iyi tanımlı hedeflerde, iç tüy hücresi sinapsının kalsiyum-duyarlı ekzositozunu onarmayı amaçlayan gen tedavileri klinik denemelere taşınır. Destek hücrelerinin yeniden programlanması (Atoh1, Notch/Wnt modülasyonu) ve vektör tasarımları, memeli kokleadaki rejenerasyon engelini aşmaya çalışır.
- Nano-ölçek yapısal biyoloji: Şeritli sinaps bileşenleri ve aday MET kanal kompleksleri, kriyo-EM ve gelişmiş görüntüleme ile daha çözünür hâle gelir; uç-bağlantı–kanal kavramı mekanik modellerle birleştirilir. Aktin çekirdeğinin kademeli mimarisi ile elastik bağlantıların frekans bağımlı davranışı, doğrudan ölçülür.
- Sistem nörobilimi ve davranış: Koklear amplifikatör kazancının beyin sapı olivokoklear demetleri ile dinamik kontrolü, dikkat ve dinleme çabası gibi bilişsel durumlarla ilişkilendirilir; tüy hücresi, artık yalnız periferik bir dönüştürücü değil, algısal ekosistemin ayarlanabilir bir düğümüdür.
11) Yan Yollarda Öğrendiklerimiz: Denge, Otoproteksiyon ve Klinik İnce Ayar
Vestibüler tarafta, otokoniyal jel ve kupulanın viskoelastikliği ile tüy demeti mekaniklerinin eşleşmesi, yaşlanma ve kalsiyum metabolizmasıyla bağlanır; benign paroksismal pozisyonel vertigonun biyofiziği anlaşılır. Otoakustik emisyon tabanlı taramalar, yeni doğan işitme kaybını erken yakalar. Ototoksisiteye –aminoglikozidler, sisplatin– karşı koruyucu stratejiler, farmakolojinin ince terazisinde tartılır; çocuk onkolojisinde duyuyu korumak artık gerçekçi bir hedef hâline gelir.
12) Dilin Kendisi: “Tüy” ve “Saç”ın Etimolojisi, Kavramın Evrimi
Türkçe’de “tüy/saç hücresi” denilen bu hücrelerin İngilizce karşılığı hair cell’dir; “saç” benzetmesi, apikal yüzeydeki stereosilya demetinin görünüşünden doğar. “Stereosilyum” Yunanca stereos (katı) ve Latince cilium (kirpik) köklerinden birleşir; vestibül (Lat. vestibulum, giriş holü) ve koklea (Yun. kochlias, salyangoz) terimleri, yapının hem biçimini hem de işlevsel topoğrafyasını dille mühürler. İlginç olan, “saç” benzetmesinin zamanla mecazdan moleküle dönüşmesidir: artık bu “saçların” aktin filamentlerinden örülü olduğu, uçlarının kadherin köprüleriyle gerilim algıladığı, açılan gözenekten potasyum ve kalsiyumun aktığı bilinir.
13) Keşiflerin Haritası: İsimler, Aletler, Kavramlar
- Malpighi – Valsalva – Cotugno: Labirentin bedenini tanımladılar.
- Corti – Deiters – Hensen – Claudius: Epitelin örgüsünü ve hücre tiplerini ayırdılar.
- Helmholtz: Frekans-seçiciliğin kuramsal iskeletini kurdu.
- Ewald – Bárány: Vestibüler fiziolojiyi deneysel ve klinik temele oturttu.
- Békésy: Seyahat eden dalgayı ölçtü; tonotopiyi kanıta çevirdi.
- Gold – Rasmussen: Aktif süreç ve efferent kontrol kavramlarını yerleştirdi.
- Engström – Wersäll: EM çağında tipoloji ve ultra-yapıyı görünür kıldı.
- Fettiplace – Corey – Hudspeth: MET akımını doğrudan kaydetti.
- Kemp: Otoakustik emisyonlarla amplifikatörü çevikçe doğruladı.
- Brownell – Zheng ve çalışma arkadaşları: OHC elektromotilitesi ve prestin ile motoru tanımladı.
- Pickles ve meslektaşları – cadherin-23/protocadherin-15: Uç-bağlantı mimarisini çözdüler.
- Genetikçiler ve sinaptik biyologlar: TMC1/TMC2, otoferlin ve onlarca sağırlık genini haritaladılar.
- House – Clark ve mühendisler: Koklear implantla klinikte yeni bir çağ açtılar.
- Rubel – Cotanche ve rejenerasyon biyolojisi: Memelilerle kuşlar arasındaki yenilenme farkını ortaya koydular.
- Klinik çeviri ekosistemi (2015–2025): Otof gen terapileri, rejenerasyon programları ve sinaptopatiyi hedefleyen stratejilerle umut veren bir eşiği yokluyor.
14) Bugünden Bakınca
Tüy hücrelerinin hikâyesi bitmiş bir öykü değil; yaşayan bir araştırma gündemidir. Bir yanda atomdan organa kadar çok katmanlı bir mekanodan elektrike dönüşüm şeması, diğer yanda klinikte ölçülebilir, tedavi edilebilir hedefler… Stereosilyanın ucundaki kadherin köprüsü gerildiğinde açılan birkaç nanometrelik gözenek, bir çocuğun ilk kez annesinin sesini işitmesine kadar uzanan bir zincirin ilk halkasıdır. Keşiflerin asıl anlamı, belki de budur.