Sinonim: Çöp DNA, Non-coding DNA, junk DNA.
Deoksiribonükleik asidin (DNA) proteinleri kodlamayan kısımlarıdır. İnsanlar, hayvanlar ve bitkiler gibi daha yüksek organizmalarda, DNA’nın büyük çoğunluğu bu anlamda “kodlamaz”. Bu kodlamayan DNA için çeşitli işlevler de bulunmuştur. İşlevli kodlamayan DNA oranının, kodlamayan DNA’nın işlevsiz oranıyla karşılaştırıldığında ne kadar büyük olduğu bilinmemektedir.

İnsan genomu yaklaşık 3,2 milyar DNA baz çifti içerir. Yine de, genomun içinde, genomun yalnızca% 1.5’ini oluşturan kabaca 20.000 protein kodlayan gen vardır. Bu genler tarafından kodlanan proteinler, hücrelerin temel bileşenleridir ve enzim, yapısal elemanlar ve sinyal molekülleri olarak işlev görür. 20.000, kodlanan proteinlerin gerçek sayısını olduğundan daha az tahmin etse de (birçok gen, farklı protein izoformlarını kodlayan çoklu RNA kopyaları üretir), yine de solucanların 1000’den az hücreden oluşması şaşırtıcıdır. Ve 30 kat daha küçük genomlarla – ayrıca kabaca 20.000 protein kodlayan genden bir araya getirilmiştir. Belki de daha da tedirgin edici olan, bu proteinlerin çoğunun insanlarda ifade edilen moleküllerin tanınabilir homologları olmasıdır. O halde insanları solucanlardan ayıran nedir?
Cevap tam olarak bilinmemektedir, ancak kanıtlar, farklılığın insan genomunun proteinleri kodlamayan% 98,5’inde olduğu iddiasını desteklemektedir. Bu kadar uzun DNA uzantılarının (genomun ‘karanlık maddesi’ olarak adlandırılan) işlevi uzun yıllar boyunca gizemliydi. Bununla birlikte, insan genomunun % 85’inden fazlasının nihayetinde kopyalanmış olduğu ve neredeyse % 80’inin gen ekspresyonunun düzenlenmesine ayrıldığı artık açıktır. Bunun sonucu olarak proteinler, hücreleri, dokuları ve organizmaları bir araya getirmek için gerekli yapı bloklarını ve makineleri sağlarken, kritik ‘mimari planlamayı’ sağlayan, genomun kodlamayan bölgeleridir. İnsan genomunda bulunan fonksiyonel protein kodlamayan DNA dizilerinin başlıca sınıfları şunları içerir :
• Protein transkripsiyon faktörlerini bağlayan promotör ve güçlendirici bölgeler
• Daha yüksek sıralı kromatin yapılarını düzenleyen ve koruyan proteinler için bağlanma yerleri
• Kodlamayan düzenleyici RNA’lar. Düzenleyici işlevlere adanmış genomun% 80’inin büyük çoğunluğu RNA’lara (mikro RNA’lar ve uzun kodlamayan RNA’lar (daha sonra açıklanacak)) dönüştürülür ve bunlar asla çevrilmez. Proteine dönüşür, ancak gen ekspresyonunu düzenleyebilir.
• Mobil genetik unsurlar (örneğin, transpozonlar). Dikkat çekici bir şekilde, insan genomunun üçte birinden fazlası bu tür ‘sıçrayan genlerden’ oluşur. Bu bölümler genom etrafında dolaşabilir ve gen düzenlemesi ve kromatin organizasyonunda rol oynar.
• Telomerler (kromozom uçları) ve sentromerler (kromozom ‘bağları’) dahil olmak üzere özel yapısal DNA bölgeleri. Önemlisi, hastalıklarla ilişkili birçok genetik varyasyon (polimorfizm), genomun protein kodlamayan bölgelerinde bulunur. Bu nedenle, jeneregülasyondaki varyasyon, spesifik proteinlerdeki yapısal değişikliklerden daha hastalık nedenselliğinde daha önemli olabilir. Genom dizilemeden ortaya çıkan bir başka sürpriz de, herhangi iki insanın tipik olarak% 99,5 DNA özdeş (ve şempanzelerle % 99 dizilim özdeş) olmasıdır! Bu nedenle, hastalıklara ve çevresel maruziyetlere karşı farklı duyarlılık dahil olmak üzere bireysel varyasyon, DNA’mızın % 0,5’inde kodlanır (daha da önemlisi, bu hala yaklaşık 15 milyon baz çiftini temsil eder).
İnsan genomundaki en yaygın iki DNA varyasyonu formu, tek nükleotid polimorfizmleri (SNP’ler) ve kopya sayısı varyasyonlarıdır (CNV’ler).
• SNP’ler, tek nükleotid pozisyonlarındaki varyantlardır ve neredeyse her zaman bialeliktir (popülasyondaki belirli bir bölgede, örneğin A veya T gibi, yalnızca iki seçenek vardır). 6 milyondan fazla insan SNP’si tanımlanmıştır ve birçoğu farklı popülasyonlarda sıklıkta geniş farklılıklar göstermektedir. Aşağıdaki özellikler dikkate alınmaya değerdir:
• SNP’ler genom boyunca – eksonlar, intronlar, genler arası bölgeler ve kodlama bölgeleri içinde meydana gelir.
• SNP’lerin kabaca % 1’i, kodlama bölgelerinde meydana gelir; bu, şans eseri beklenecek olanla ilgilidir, çünkü kodlama bölgeleri genomun yaklaşık % 1.5’ini oluşturur.
• Kodlamayan bölgelerde bulunan SNP’ler, genomdaki düzenleyici elemanlarda meydana gelebilir ve dolayısıyla gen ekspresyonunu değiştirebilir; Bu gibi durumlarda SNP, hastalığa yatkınlık üzerinde doğrudan bir etkiye sahip olabilir.
• SNP’ler ayrıca gen fonksiyonu veya taşıyıcı fenotipi üzerinde hiçbir etkisi olmayan ‘nötr’ varyantlar olabilir.

• “Nötr” SNP’ler bile, fiziksel yakınlığın bir sonucu olarak hastalıkla ilişkili bir genle birlikte miras alınırlarsa yararlı belirteçler olabilir. Başka bir deyişle, SNP ve nedensel genetik faktör bağlantı içindedir
Dengesizlik.
• Çoğu SNP’nin hastalığa yatkınlık üzerindeki etkisi zayıftır ve bu tür varyantların tanımlanmasının, tek başına veya kombinasyon halinde, hastalık tahmini veya önleme için etkili stratejiler geliştirmek için kullanılıp kullanılamayacağı görülecek.
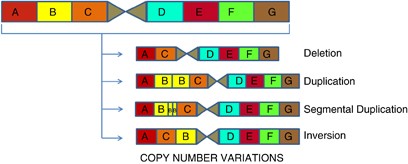
• CNV’ler, farklı sayılarda büyük bitişik DNA uzantılarından oluşan bir genetik varyasyon biçimidir; Bunlar 1000 baz çiftinden milyonlarca baz çiftine kadar değişebilir. Bazı durumlarda, bu lokuslar, SNP’ler gibi, bialeliktir ve popülasyonun bir alt kümesinde basitçe kopyalanır veya silinir. Diğer durumlarda, insan popülasyonunda çok sayıda alel bulunan, genomik materyalin karmaşık yeniden düzenlemeleri vardır. CNV’ler birkaç taneden sorumludur.
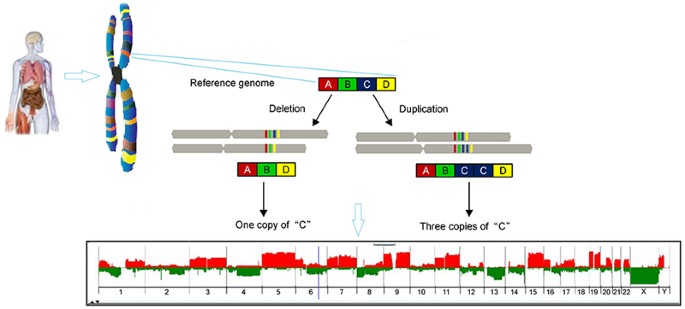
CNV’lerin yaklaşık % 50’si gen kodlama dizilerini içerir; bu nedenle CNV’ler, insan fenotipik çeşitliliğinin büyük bir kısmının altında yatabilir.
DNA sekansındaki değişikliklerin kendi başlarına insan popülasyonlarındaki fenotip çeşitliliğini açıklayamayacağına dikkat etmek önemlidir; dahası, klasik genetik kalıtım, monozigotik ikizlerdeki farklı fenotipleri açıklayamaz. Bu bilmecelerin cevapları muhtemelen epigenetikte yatmaktadır – DNA dizisindeki değişikliklerden kaynaklanmayan gen ifadesinde kalıtsal değişiklikler ayrıca bakınız.