Kompaksiyon
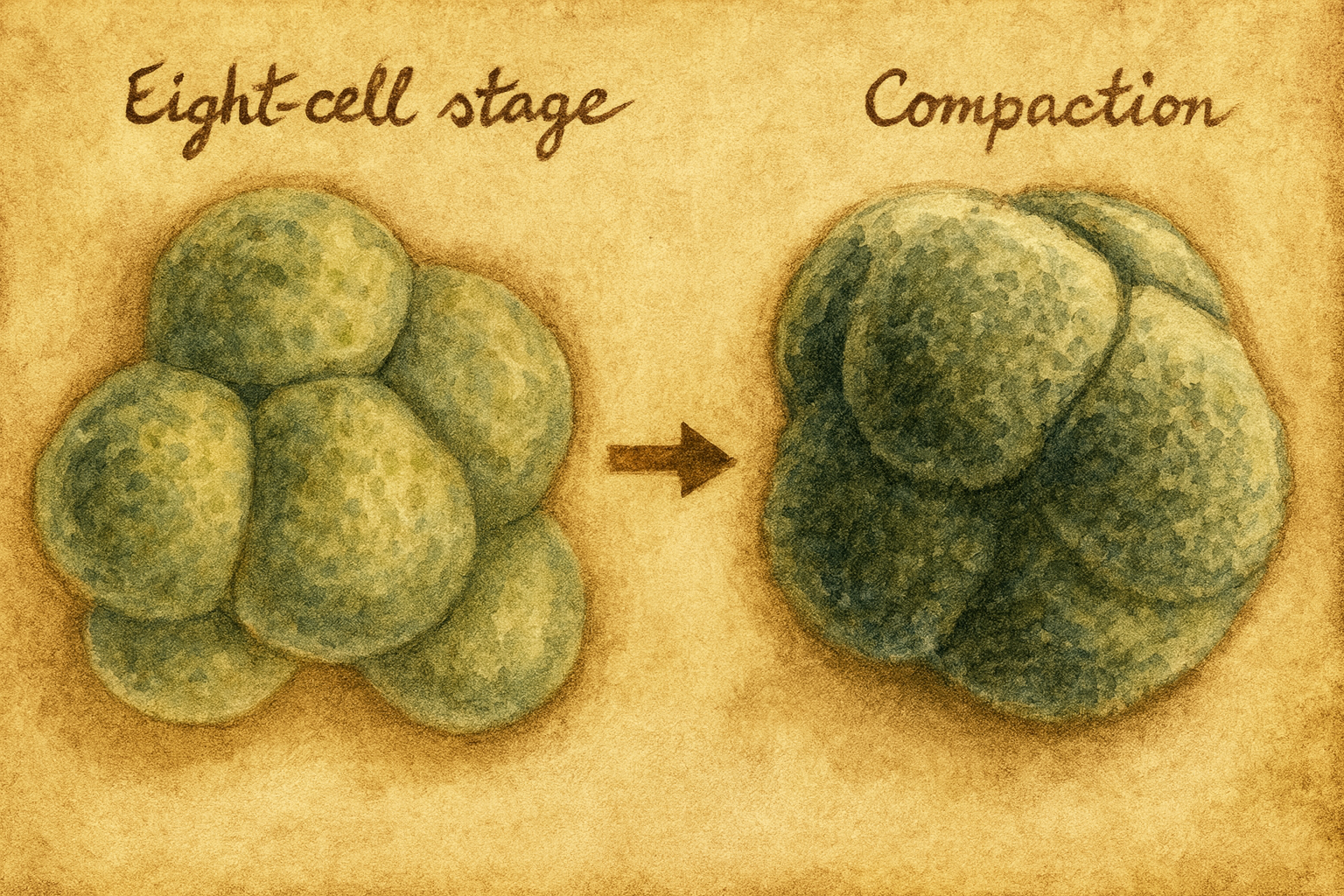
Etymoloji ve Kavramsal Evrim Latince compāctiō/compactionem “bir araya getirme, sıkılaştırma” anlamına gelir ve com- (“birlikte”) öneki ile pangere (“tutturmak, bağlamak, sabitlemek”) fiil kökünün geçmiş ortaç biçimi (compāctus) üzerinden türemiştir. Orta Çağ Latincesinden Eski Fransızcaya geçerken “yapma eylemi veya kompakt olma durumu” anlamı korunmuş, İngilizcede 14. yüzyıl sonlarından itibaren “sıkılaşma/sıkıştırma” karşılığında yerleşmiştir. Embriyolojide “kompaksiyon” terimi, morula … Devamını oku